Общая Физиология 1 Аналитический и системный подход к изучению функций организма При аналитическом подходе
 Скачать 1.8 Mb. Скачать 1.8 Mb.
|

|
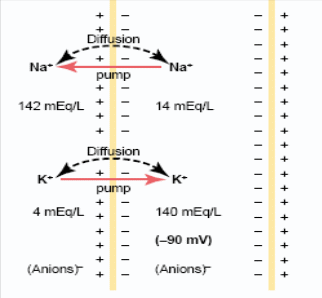
№7 Мембранно-ионная теория возбуждения,раздражимость и возбудимость как основа реагирования организма на воздействия РАЗДРАЖИМОСТЬ – как основное свойство живого. Все живые ткани отвечают на действие раздражителя изменением своего состояния – раздражимостью, что является очень широким биологическим понятием, а по сути – это ответная биологическая реакция. виды раздражимости: Неспецифическая – присуща всем живым структурам. Проявляется через способность клетки реагировать на самые разнообразные раздражения. Специфическая, или избирательная – появляется в способности определенных структур реагировать на определенные виды раздражителей (кванты света и т.д.). В гипоталамусе – осморецепторы, рецепторы на концентрацию углеводов в крови, аминокислот и т.д. Триггерная или курковая – это высший вид специфической раздражимости – возбудимость. Это свойство присуще только высокоорганизованным тканям – мышечной, нервной, железистой. Возбудимость– это способность отвечать на раздражение возбуждением, т.е. совокупностью изменений физических, химических и функциональных. Наиболее типичным проявлением процесса возбуждения является изменение электрического состояния ткани. В состоянии физиологического покая регистрируется разность потенциалов на мембране клеток – потенциал покоя (ПП), при возбуждении ПД. Мембранно-ионная теория Согласно мембранно-ионная теория, наличие электрических потенциалов в живых клетках обусловлено неравенством концентраций ионов Na+, K+, Ca2+ и Cl- внутри и вне клетки. И различной проницаемостью для них поверхностной мембраны, а именно: Клеточная мембрана обладает селективной (избирательной) проницаемостью для ионов: в состоянии покоя для ионов К+ в 25 раз больше, чем для Na+, а в состоянии возбуждения – натриевая проницаемость в 20 раз больше калиевой. Концентрация ионов Na+, K+ и Cl- различна внутри и вне клетки: Движение ионов через воротную ситему мембраны осуществляется двумя механизмами: свободной диффузией по градиенту концентрации (пассивный транспорт) и транспортной системой − Na+,K+-насосом – молекулярным механизмом, локализованным в мембране, которая представляет собой белковую молекулу, способную транспортировать вещества, используя энергию распада АТФ под действием АТФазы. В состоянии покоя Na+,K+-насос переносит 3 иона Na+ на наружную поверхность мембраны против градиента концентрации, а в обратном направлении во внутрь клетки 2 иона К+ также против градиента концентрации. Таким образом, данный механизм имеет значение: а) создает и поддерживает трансмембранный градиент концентрации для ионов Na+ и К+ в состоянии покоя и в состоянии возбуждения; б) формирует разность потенциалов, суммирующуюся с потенциалом покоя или потенциалом действия. Это происходит потому, что насос электрогенен: на 3 иона Na+, вынесенного из клетки, вносится 2 иона К+, т.е. 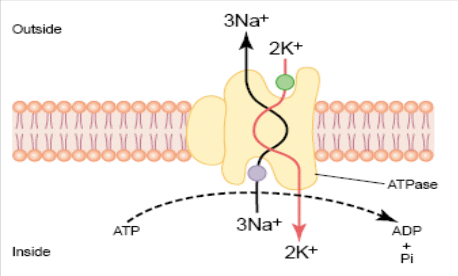 Указанные процессы приводят к формированию электрического состояния ткани как в покое, так и в возбуждении. №8 Физиологическая сущность потенциала покоя; его происхождение ПОТЕНЦИАЛ ПОКОЯ Термином мембранный потенциал или ПП принято называть трансмембранную разность потенциалов, существующую между цитоплазмой и окружающей средой (наружным раствором). Когда клетка находится в состоянии физиологического покоя, её внутренний потенциал электроотрицателен по отношению к наружному. У различных клеток потенциал варьирует от −50 до −90 мВ. Измерение ПП: техника внутриклеточных микроэлектродов. Природа ПП. В состоянии физиологического покоя мембрана больше проницаема (в 25 раз) для ионов К+, чем для ионов Na+ и Cl-. Следовательно, К+ устремляется из клетки по градиенту концентрации в наружную среду. Большие органические анионы не могут пройти через мембрану и поэтому не способны сопровождать ионы К+. В этом случае каждый выходящий ион К+ будет оставлять после себя в клетке отрицательный заряд. Такое движение ионов К+ по законам диффузии будет происходить до тех пор, пока их концентрация по обе стороны мембраны не выровняется. Но разности потенциалов в этом случае ещё сформироваться не может. Подключается механизм ионного насоса, который постоянно поддерживает градиент концентрации ионов К+ и, таким образом, обеспечивает постоянное движение ионов К+ через систему ионных каналов мембраны клетки. В результате между внутренней и внешней поверхностями клетки возникает разность потенциалов, которая, если учитывать только движение ионов К+, согласно  Однако мембрана проницаема и для ионов Na+ и Cl-, хотя очень незначительно. Это снижает величину ПП до −80 −70 мВ. Как это происходит, наглядно демонстрирует формула Ходжкина-Катца:  , ,где учитывается не только концентрация всех потенциалобразующих ионов, но и проницаемость мембраны для них в данном состоянии клетки. В результате всего описанного, мембрана в покое находится в состоянии статической (постоянной) поляризации, внутри клетка заряжается элеткроотрицательно по отношению к наружной поверхности мембраны.
При нанесении на клетку мышечной ткани или нервное волокно порогового раздражения мембранный потенциал начинает изменяться, что называется состоянием деполяризации, в результате которой происходит перезарядка мембраны и формирование ПД – потенциала действия, который отражает наличие в клетке или нервном волокне процесса возбуждения. №9 Современное представления о формировании потенциала действия ПОТЕНЦИАЛ ДЕЙСТВИЯ Причиной возникновения потенциала действия – ПД – в нервных и мышечных волокнах является изменение ионной проницаемости их мембраны. При действии на клетку раздражителя проницаемость мембраны для ионов Na резко увеличивается (PNa в 20 раз больше, чем РК). Поэтому поток положительно заряженных ионов Na внутрь клетки начинает значительно превышать поток ионов К из клетки. Происходит это движение по механизму пассивного транспорта, выравниванию же концентрации ионов Na по обе стороны мембраны препятствует, как и в случае с ПП, работа Na,К-насоса. В результате этого происходит перезарядка мембраны – её деполяризация. Плотность ионных каналов на поверхности мембраны: для ионов Na 50 на 1 мкм2 для ионов К несколько меньше Величина пор: для ионов Na → 0,8 нм для ионов К → 0,3 нм В результате описанных процессов возникает разность потенциалов обратного знака: внутреннее содержимое клетки становится заряженным положительно по отношению к наружной среде, иными словами, происходит реверсия мембранного потенциала до величины +30-+40 мВ. Фазы потенциала действия: ав – местная деполяризация (локальный ответ), вс – распространяющаяся деполяризация, восходящая часть «спайк»-потенциала, сd – реполяризация (нисходящая часть «спайк»-потенциала), de – следовая деполяризация (следовой отрицательный потенциал), ef – следовая деполяризация (следовой положительный потенциал). Таким образом, в состоянии возбуждения на мембране развивается:
Процесс деполяризации связан с селективной проницаемостью ионных каналов для Na. Повышение проницаемости для ионов Na продолжается очень короткое время – 1 мс! Вслед за этим в клетке начинаются восстановительные процессы, приводящие к тому, что проницаемость для Na вновь уменьшается, а для К – возрастает. Это называется натриевой инактивацией. В результате поток положительно заряженных ионов Na внутрь протоплазмы резко ослабевает, а увеличение калиевой проницаемости вызывает усиление потока положительно заряженных ионов К из протоплазмы во внешний раствор. Параллельно усиливается работа Na,К–насоса по возвращению к исходной полярности. Это приводит к реполяризации мембраны, т.е. возвращению к исходной полярности. Состояние гиперполяризации развивается в результате повышенной калиевой проницаемости мембраны в конце процесса возбуждения, которая возникает как «шлейф» последействия процесса возбуждения. В результате мембрана получает разность потенциалов еще большую, чем была в состоянии покоя. Соответствие фаз потенциала действия электрическим состояниям мембраны Деполяризация – происходит перезарядка мембраны сначала до нуля, а затем, так как поток ионов Na во внутрь клетки продолжается, формируется заряд на поверхности мембраны +30 мВ (АВ – местный потенциал, ВС – восходящая ветвь спайк-потенциала). Реполяризация – натриваевая инактивация приводит к снижению потока Na внутрь клетки, калиевая проницаемость растет, полярность клетки стремится к исходной величине до -60 мВ (CD – нисходящая часть спайк-потенциала). Реполяризация происходит волнообразно, что отражается через формирование следового отрицательного потенциала – тормозится калиевая проницаемость, а проницаемость для Na растет. Формируется разность потенциалов – СОП, который затем становится равным ПП - -80 мВ (отрезок EF). Гиперполяризация – количественно выражается следовым положительным потенциалом – до -100 мВ – СПП (отрезок FK). Следовые потенциалы. Следовые потенциалы являются очень чувствительными к концентрации ионов К+ во внеклеточной среде: при увеличении содержания К+ они уменьшаются. Следовые потенциалы впервые были зарегистрированы Д.С. Воронцовым в 1926 г., а затем подробно изучены Эрлангером и Гассером. Амплитуда этих потенциалов не превышает нескольких милливольт, а длительность – от нескольких миллисекунд до нескольких десятков и сотен секунд. Обусловлены следовые потенциалы восстановительными процессами в клетке вслед за возбуждением. №10 Физиологическая сущность местного и распространяющегося возбуждения Возбуждение может быть 2-х видов: местное (локальный ответ); распространяющееся (импульсное). Местное возбуждение- наиболее древний вид (низшие формы организмов и низковозбудимые ткани - например, соединительная ткань). Местное возбуждение возникает и в высокоорганизованных тканях под действием подпорогового раздражителя или как компонент потенциала действия. При местном возбуждении нет видимой ответной реакции. Особенности местного возбуждения: нет латентного (скрытого) периода - возникает сразу же при действии раздражителя; нет порога раздражения; местное возбуждение градуально - изменение заряда клеточной мембраны пропорционально силе подпорогового раздражителя; нет рефрактерного периода, наоборот характерно небольшое повышение возбудимости; распространяется с декрементом (затуханием). Импульсное (распространяющееся) возбуждение- присуще высокоорганизменным тканям, возникает под действием порогового и сверхпорогового раздражителей. Особенности импульсного возбуждения: имеет латентный период - между моментом нанесения раздражения и видимой ответной реакцией проходит некоторое время; имеет порог раздражения; не градуально - изменение заряда клеточной мембраны не зависит от силы раздражителя; наличие рефрактерного периода; импульсное возбуждение не затухает. Вывод: в организме животного и человека наблюдается местное и импульсное возбуждение. Возникновение того или иного вида возбуждения зависит от степени развития ткани и силы раздражителя. №11 Динамика изменения возбудимости в процессе формирования потенциала действия. Соотношение фаз возбудимости и потенциала действия. Рефрактерность:её физиологическая сущность Способность формировать новый процесс возбуждения в условиях частотного раздражения зависит от изменения возбудимости в процессе возбуждения. Для выявления этого на возбудимую ткань наносят два последовательно действующих раздражителя в интервале времени, соответствующем: 1) ЛО – МП (локальному ответу – местному потенциалу); 2) Спайк-потенциалу (его восходящей и нисходящей ветви) 3) СОП – следовый отрицательный потенциал; 4) СПП – следовый положительный потенциал; В результате выяснилось: 1. Во время локального ответа (местной деполяризации) возбудимость несколько повышена – достаточно действия допорогового раздражителя для возникновения «спайк-потенциала». 2. Периоду возникновения и развития пика «спайк-потенциала» соответствует полная невозбудимость клетки –абсолютная рефрактерность. Ткань не реагирует даже на сверхпороговые раздражители. Длительность этой фазы совпадает с длительностью спайка ПД – 0,4-0,5 мс. 3. В условиях реполяризации возбудимость постепенно восстанавливается и ткань может отреагировать, но только на сверхпороговые раздражители – относительная рефрактерность. Согласно Ходжкину и Хаксли рефрактерность обусловлена тем, что вслед за начальной деполяризацией, приводящей к возникновению потенциала действия, развивается процесс натриевой инактивации, и в то же время повышается калиевая проницаемость для мембраны. Результатом этого является невозможность мембраны отвечать потенциалом действия на новое раздражение. Постепенно устраняется натриевая инактивация и нормализуется калиевая – формируется состояние относительной рефрактерности. В конце фазы относительной рефрактерности возбудимость не только восстанавливается до нормального уровня, но и превышает его. Это фаза супернормальной возбудимости (ткань может отреагировать на подпороговый раздражитель). Механизм заключается в резком снижении критического уровня деполяризации. Чем больше длится следовая деполяризация, тем дольше фаза супернормальной возбудимости (средняя длительность – 12-30 мс). В тех тканях, где отмечается гиперполяризация – СПП - за фазой супернормальной возбудимости следует фаза субнормальной возбудимости. Механизм снижения возбудимости в эту фазу заключается в увеличении критического уровня деполяризации. Д 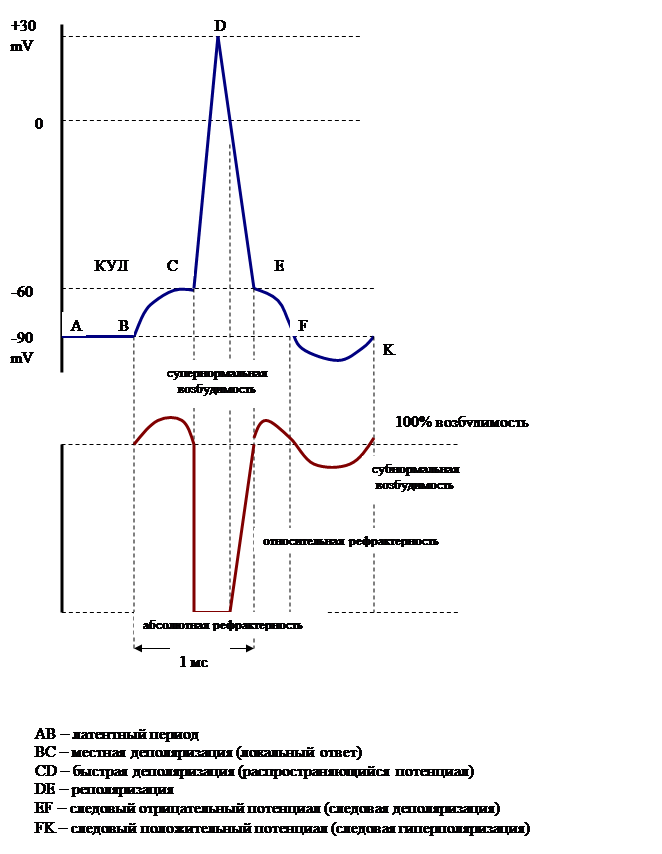 ИНАМИКА ВОЗБУДИМОСТИ ПРИ ВОЗБУЖДЕНИИ Рефрактерность – это неспособность клетки воспринимать нервный импульс, что проявляется в отсутствии возбудимости при действии раздражителя вследствие изменения состояния потенциалзависимых каналов в мембране. Выделяют периоды абсолютной и относительной рефрактерности. их продолжительность зависит от продолжительности ПД. Период абсолютной рефрактерности, характеризующееся отсутствием ответа на действие любой силы раздражителя, является следствием инактивации Na +-каналов, открытие которых необходимо для возникновения нового ПД. Инактивация длится около 0,25 с. Натриевый ток начинает восстанавливаться только после того, как мембраны кардиомиоцитов реполяризувалися примерно до уровня - 40 мВ. В период относительной рефрактерности действие сильного, чем обычно, раздражителя может вызвать появление внеочередного ПД. Период относительной рефрактерности продолжается 0,03 с. После него в течение некоторого времени, наблюдается супернормальная возбудимость - когда сердечная мышца может возбуждаться также под влиянием подпорогового раздражителя. Для миокарда продолжительность периода рефрактерности имеет особое значение. Она предотвращает слишком частых повторных возбуждений миокарда. Такие возбуждения в скелетных мышцах приводят к тетануса. Но подобный ответ со стороны миокарда сделала бы невозможной нагнетательную функцию сердца. При рефрактерности желудочки успевают расслабиться и начинают заполняться кровью. Кроме того, рефрактерность «не позволяет» возбуждению бесконечно долго циркулировать по миокарда. Начавшись в предсердиях, оно снова могло бы вернуться к ним, обойдя желудочки. Но в это время предсердия находятся в стадии полной рефрактерности, а это значит, что новая волна возбуждения возникнуть в них не может. №12 Законы раздражения возбудимых тканей: Гоорвега-Вейса-Лапика, Пфлюгера, Боудича,Дюбуа-Реймона 1) Зависимость пороговой силы стимула от его длительности.- Закон Гоорвега-вейса-Лапика Пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. Эта зависимость, открытая Гоорвегом, Вейсом, Лапиком получила название кривой “сила – длительность” или “сила – время”. Кривая “силы – времени” имеет форму, близкую к равносторонней гиперболе и в первом приближении может быть описана эмпирической формулой: I= a + b , где I – сила тока T T – длительность его действия а, b – постоянные, определяемые свойствами ткани. Из этой кривой следует: Ток величиной ниже порога не вызывает возбуждения, как бы длительно он не действовал. Какой бы сильный не был раздражитель, но если он действует очень короткое время, то возбуждение не возникает. Минимальная сила тока (или напряжение), способная вызвать возбуждение, названа реобазой – (основание тока)=порогу. Наименьшее время, в течение которого должен действовать стимул в одну реобазу, чтобы вызвать возбуждение – полезное время. Дальнейшее его увеличение не имеет значения для возникновения возбуждения. Порог (реобаза) – величины непостоянные, зависят от функционального состояния клеток в покое. Поэтому Лапик предложил определять более точный показатель – хронаксию. Хронаксия – наименьшее время, в течение которого ток в две реобазы должен действовать на ткань, чтобы вызывать возбуждение. Определение хронаксии – хронаксиметрия – получило распространение в клинике для диагностики повреждения нервных стволов и мышц 2)Полярный закон раздражения (закон Пфлюгера). (изменения мембранного потенциала при действии на возбудимые ткани постоянного электрического тока). Пфлюгер (1859) Постоянный ток проявляет свое раздражающее действие только в момент замыкания и размыкания цепи. При замыкании цепи постоянного тока возбуждение возникает под катодом; при размыкании по анодом. Изменение возбудимости под катодом. При замыкании цепи постоянного тока под катодом (действуют допороговым, но продолжительным раздражителем) на мембране возникает стойкая длительная деполяризация, которая не связана с изменением ионной проницаемости мембраны, а обусловлена перераспределением ионов снаружи (привносятся на электроде) и внутри – катион перемещается к катоду. Вместе со смещением мембранного потенциала смещается и уровень критической деполяризации – к нулю. При размыкании цепи постоянного тока под катодом мембранный потенциал быстро возвращается к исходному уровню, а УКД медленно, следовательно, порог увеличивается, возбудимость снижается – катодическая депрессия Вериго. Таким образом, ввозникает только при замыкании цепи постоянного тока под катодом. Изменение возбудимости под анодом. При замыкании цепи постоянного тока под анодом (допороговый, продолжительный раздражитель) на мембране развивается гиперполяризация за счет перераспределения ионов по обе стороны мембраны (без изменения ионной проницаемости мембраны) и возникающее за ней смещение уровня критической деполяризации в сторону мембранного потенциала. Следовательно, порог уменьшается, возбудимость повышается – анодическая экзальтация. При размыкании цепи мембранный потенциал быстро восстанавливается к исходному уровню и достигает сниженного уровня критической деполяризации, генерируется потенциал действия. Таким образом, возбуждение возникает только при размыкании цепи постоянного тока под анодом. Сдвиги мембранного потенциала вблизи полюсов постоянного тока получили название электротонических. Сдвиги мембранного потенциала не связанные с изменением ионной проницаемости мембраны клетки называют пассивными 3) Закон “ все или ничего”.-Закон Боудича Установлен Боудичем в 1871 г. на мышце сердца. При подпороговой силе раздражения мышца сердца не сокращается, а при пороговой силе раздражения – сокращение максимально. При дальнейшем увеличении силы раздражения амплитуда сокращений не увеличивается. Со временем была установлена и относительность этого закона. Оказалось, что “все” зависит от функционального состояния ткани (охлаждение, исходное растяжение мышцы и т. д.). С появлением микроэлектродной техники было установлено еще одно несоответствие: подпороговое раздражение вызывает местное, нераспространяющееся возбуждение, следовательно, нельзя говорить, что допороговое раздражение не дает ничего. Процесс развития возбуждения подчиняется этому закону с уровня критической деполяризации, когда запускается лавинообразное поступление ионов калия в клетку. 4)Закон раздражения Дюбуа-Реймона (аккомодации): раздражающее действие постоянного тока зависит не только от абсолютной величины силы тока или его плотности, но и от скорости нарастания тока во времени. При действии медленно нарастающего раздражителя возбуждение не возникает, так как происходит приспосабливание возбудимой ткани к действию этого раздражителя, что получило название аккомодации.Аккомодация обусловлена тем, что при действии медленно нарастающего раздражителя в мембране возбудимой ткани происходит повышение критического уровня деполяризации. При снижении скорости нарастания силы раздражителя до некоторого минимального значения потенциал действия вообще не возникает. Причина заключается в том, что деполяризация мембраны является пусковым стимулом к началу двух процессов: быстрого, ведущего к повышению натриевой проницаемости, и тем самым обусловливающего возникновение потенциала действия, и медленного, приводящего к инактивации натриевой проницаемости и как следствие этого - окончанию потенциала действия. При быстром нарастании стимула повышение натриевой проницаемости успевает достичь значительной величины прежде, чем наступит инактивация натриевой проницаемости. При медленном нарастании тока на первый план выступают процессы инактивации, приводящие к повышению порога или ликвидации возможности генерировать ПД вообще. Способность к аккомодации различных структур неодинакова. Наиболее .высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника, желудка №13 Физиологическая сущность понятий: функция, физиологическая система, живой организм 1.Функция – это специфическая деятельность клеток, органов и систем органов по обеспечению жизнедеятельности целого организма. Это проявления жизнедеятельности, имеющие приспособительное значение, осуществляя различные функции, организм приспособляется к внешней среде или же приспособляет среду к своим потребностям. 2)Физиологическая система - совокупность органов и тканей, выполняющих одну функцию (костная, мышечная ит. п. ) 3)Живой организм"означает любое биологическое образование,которое способно к передаче или репликации генетического материала, включая стерильные организмы,вирусы и вироиды №14 Функциональное единство организма с окружающей средой Все системы органов находятся в сложном взаимодействии друг с другом и составляют в генетическом, анатомическом и функциональном отношении единое целое - организм. Только благодаря своей целостности организм обладает совокупностью основных жизненных свойств, отличающих его от неживой материи. К ним относятся: 1) постоянный обмен веществ и энергии с окружающей средой и внутри самого организма, характеризующийся самообновлением; 2) движение; 3) рост и развитие; 4) размножение; 5) изменчивость; 6) наследственность; 7) приспособляемость к условиям существования. Саморазвитие, самовоспроизведение и самоуправление, являясь основными свойствами организма как целостной биологической системы, обеспечиваются внутренними процессами диалектического взаимодействия между частями и целым, между целым (организмом) и окружающей его средой. Целостность организма обеспечивается как механически, путем соединения клеток, тканей, органов и др. в единую массу, так и функционально (нейро-гуморальная регуляция). На организм человека постоянно воздействует природная и социальная среда. Разнообразные факторы внешней среды действуют на организм через органы чувств и нервную систему, которая, объединяя в единое целое организм, регулирует деятельность его внутренней среды, приспосабливает обмен веществ к меняющимся условиям жизни, уравновешивает организм с окружающей его средой. Единство организма и внешней среды составляет основу эволюции животных организмов. Важнейшим фактором социальной среды, воздействующим на человека, является труд. Трудовые процессы обеспечиваются специальной работой нервной и мышечной систем. Вместе с тем трудовая (умственная и физическая) деятельность человека направлена на преобразование природы и ее приспособление к нуждам человека. На организм человека оказывают существенное влияние такие социальные факторы, как условия труда и быта, питание и одежда, режим труда и отдыха, удовлетворение духовных потребностей и т. п. Поэтому большое значение имеет социальное положение человека, классовая структура общества. Физиология нервно-мышечной системы №1 Физиологический особенности скелетной мышцы. Современное представление о моторных единицах: классификация, структурно-функциональная характеристика и значение. Строение мышцы. Скелетная мышца представляет собой сложную систему, преобразующую химическую энергию в механическую работу и тепло. В настоящее время хорошо исследованы молекулярные механизмы этого преобразования. Мышечное волокно является многоядерной структурой, окруженной мембраной и содержащей специализированный сократительный аппарат — миофибриллы [1]. Кроме этого, важнейшими компонентами мышечного волокна являются митохондрии, системы продольных трубочек — саркоплазматическая сеть (ретикулум) и система поперечных трубочек — Т-система. Функциональной единицей сократительного аппарата мышечной клетки является саркомер, из саркомеров состоит миофибрилла Саркомеры отделяются друг от друга Z-пластинками. Саркомеры в миофибрилле расположены последовательно, поэтому сокращение саркомеров вызывает сокращение миофибриллы и общее укорочение мышечного волокна Мышца состоит из пучков поперечнополосатых мышечных волокон, соединенных рыхлой соединительной тканьюв пучки первого порядка. Они, в свою очередь, объединяются в пучки второго порядка и т. д. В итоге мышечные пучки всех порядков объединяются соединительной оболочкой, образуя мышечное брюшко. Соединительнотканные прослойки, имеющиеся между мышечными пучками по концам брюшка, переходят в сухожильнуючасть мышцы, крепящейся к кости. Вовремя сокращения происходит укорочение мышечного брюшка и сближение ее концов. При этом сократившаяся мышца с помощью сухожилия тянет за собой кость, которая выполняет роль рычага. Так совершаются разнообразные движения. Каждая мышцз является целостным (отдельным) органом, имеющим определенную форму, строение и функцию, развитие и положение в организме. Мышцы обильно снабжены кровеносными сосудами и нервами. В каждом движении принимают участие несколько мышц. Мышцы, действующие совместно в одном направлении и вызывающие сходный эффект, называются синергистами, а совершающие противоположно направленные движения —антагонистами. Например, сгибателем локтевого сустава является двуглавая мышца плеча (бицепс), а разгибателем — трехглавая (трицепс)- Сокращение мышц-сгибателей локтевого сустава сопровождается расслаблением мышц-разгибателей. Однако при постоянной нагрузке на сустав (например, при удержании гири в горизонтально вытянутой руке) мышцы-сгибатели и разгибатели локтевого сустава действуют уже не как антагонисты, а как синергисты. Таким образом, действия мышц нельзя сводить к выполнению только одной функции, так как они многофункциональны. Поскольку в каждом движении участвуют мышцы как одной, так и другой группы, наши движения точны и плавны. По характеру выполняемых основных движений и по действию на сустав различают следующие виды мышц: сгибатели и разгибатели, приводящие и отводящие, вращающие, приподнимающие и опускающие и др. Выделяют также мимические, жевательные и дыхательные мышцы Моторные единицы: Группа мышечных волокон и иннервирующий их мотонейрон определяются как моторная единица -минимальная функциональная единица мышечной системы. Число мышечных волокон, составляющих моторную единицу варьирует от мышцы к мышце. Там, где требуется тонкий контроль движений, например, в пальцах или мышцах глаз, моторные единицы малы, как правило не более 5 - 20 волокон; там, где тонкий контроль не требуется - в мышцах спины и бедра, моторные единицы намного крупнее и состоят из более, чем тысячи волокон. Моторные единицы одной мышцы могут быть разными. В зависимости от скорости сокращения моторные единицы разделяют на медленные/slow (S-МЕ) и быстрые/fast (F-МЕ). А F-МЕ в свою очередь делят по устойчивости к утомлению на быстроутомляемые/fast-fatigable (FF-МЕ) и устойчивые к утомлению/fast-fatigue-resistant (FR-МЕ). Классификация и значение: Соответствующим образом подразделяют иннервирующие данные МЕ мотонейроны. Существуют S-мотонейроны(S-МН), FF-мотонейроны (F-МН) и FR -мотонейроны(FR-МН) S- МЕ характеризуются высоким содержанием белка миоглобина, который способен связывать кислород (О2). Мышцы, преимущественно состоящие из МЕ этого типа, за их темно-красный цвет называются красными. Красные мышцы выполняют функцию поддержания позы человека. Предельное утомление таких мышц наступает очень медленно, а восстановление функций происходит наоборот, очень быстро. Такая способность обуславливается наличием миоглобина и большого числа митохондрий. МЕ красных мышц, как правило, содержат большое количество мышечных волокон. FF-МЕ составляют мышцы, способные выполнять быстрые сокращения без заметного утомления. Волокна FF-ME содержат большое количество митохондрий и способны образовывать АТФ путём окислительного фосфорилирования. Как правило, число волокон в FF-ME меньше, чем в S-ME. Волокна FR-ME характеризуются меньшим содержанием митохондрий, чем в FF-ME, а также тем, что АТФ в них образуется за счет гликолиза. В них отсутствует миоглобин, поэтому мышцы, состоящие из МЕ этого типа, называют белыми. Белые мышцы развивают сильное и быстрое сокращение, но довольно быстро утомляются. №2 Виды и режимы сокращения скелетных мышц. Анализ одиночного цикла сокращения-расслабления мышцы; его периоды Виды: Выделяют одиночные и тетанические виды сокращения мышц. 1. Одиночное сокращение мышцы– это сокращение, которое возникает при действии на мышцу одиночного электрического или нервного импульса. На кривой одиночного сокращения выделяют три основных фазы: 1. латентный период – время нанесения раздражения до начала сокращения. В этот период возникает возбуждение мышечных волокон и его распространение вдоль мембраны. Продолжительность варьирует до 10 мс; 2. фаза укорочения (или развитие напряжения) – около 50мс; 3. фаза расслабления - около 60 мс.  2. Тетаническое сокращение – это длительное укорочение мышцы, возникающее под действием ритмического раздражения. В его основе лежит суммация одиночных сокращений. При тетаническом сокращении амплитуда больше, чем при одиночном сокращении, так как повторные потенциалы действия возникают прежде, чем саркоплазматический ретикулум сможет удалить ранее высвобожденный кальций, поэтому уровень последнего в гиалоплазме повышается, активное состояние продлевается, увеличивается количество работающих мостиков и, как результат, усиливается сила сокращения. Для возникновения тетануса необходимо, чтобы интервал между стимулами был больше рефрактерного периода, но короче всей длительности сократительного ответа. Тетаническое сокращения имеет два вида: зубчатый тетанус и гладкий. Если повторить раздражение в фазу расслабления, то получится зубчатыйтетанус, если же в фазу укорочения – то гладкий. При некоторой достаточно высокой частоте раздражения нерва амплитуда гладкого тетануса становится наибольшей. Такой гладкий тетанус называется оптимумом. Для развития оптимума необходимо, чтобы повторные раздражители поступали к мышце после завершения периода рефрактерности, вызванного предыдущим раздражителем. Если повышать дальше частоту раздражения, то наступает состояние, которое называется пессимумом Введенского – формируется блок проведения возбуждения в нервно-мышечном синапсе и мышца вместо того, чтобы продолжать возбуждаться, расслабляется, сколько бы мы её не раздражали. В естественных условиях ввиду асинхронности работы мотонейронов сокращение мышцы напоминает гладкий тетанус. Режимы: В зависимости от условий (величины) нагрузки), при которых происходит мышечное сокращение, различают три его основных режима: 1. Изотонический режим – это сокращение мышцы, при котором её волокна укорачиваются, но напряжение остается постоянным. В реальных условиях чисто изотоническое сокращение отсутствует. 2.Изометрический режим – сокращение мышцы, при котором её длина не изменяется, но развиваемое ею напряжение возрастает. Например, поднятие груза, который больше силы мышцы. 3. Ауксотонический режим – это сокращение мышцы, при котором одновременно изменяется длина и напряжение. Этот режим характерен для натуральных двигательных актов. Одиночное мышечное сокращение. При раздражении мышцы одиночным импульсом тока возникает одиночное мышечное сокращение. Амплитуда одиночного сокращения мышцы зависит от количества сократившихся в этот момент миофибрилл. Возбудимость отдельных групп волокон различна, поэтому пороговая сила тока вызывает сокращение лишь наиболее возбудимых мышечных волокон. Амплитуда такого сокращения минимальна. При увеличении силы раздражающего тока в процесс возбуждения вовлекаются и менее возбудимые группы мышечных волокон; амплитуда сокращений суммируется и растет до тех пор, пока в мышце не останется волокон, не охваченных процессом возбуждения. В этом случае регистрируется максимальная амплитуда сокращения, которая не увеличивается, несмотря на дальнейшее нарастание силы раздражающего тока. Фазы и режимы сокращения скелетной мышцы Фазы мышечного сокращения При раздражении скелетной мышцы одиночным импульсом электрического тока сверхпороговой силы возникает одиночное мышечное сокращение, в котором различают 3 фазы (рис. 4, А): • латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения; возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия; • фаза укорочения (около 50 мс); • фаза расслабления (около 50 мс). №3 Физиологическая сущность тетануса. Оптимум и пессимум частоты стимуляции (Н.Е. Введенский) Тетанус, тетаническое мышечное сокращение (др.-греч. τέτανος — оцепенение, судорога) — состояние длительного сокращения, непрерывного напряжения мышцы, возникающее при поступлении к ней через мотонейрон нервных импульсов с высокой частотой. При этом расслабления между последовательными одиночными сокращениями не происходит и возникает их суммация, приводящая к стойкому максимальному сокращению мышцы. Различают зубчатый и гладкий тетанус. При зубчатом тетанусе каждый последующий нервный импульс воздействует на начавшую расслабляться мышцу, при этом происходит неполная суммация сокращений. При гладком тетанусе, имеющем бо́льшую амплитуду, воздействие импульса происходит в конце периода укорочения, что приводит к полной суммации сокращений Н. Е. Введенский (1886) установил, что возбуждение и торможение — фазы единого нервного процесса, которые при определенных условиях переходят друг в друга. Переход возбуждения в торможение, и наоборот, зависит от частоты и силы раздражения и от уровня лабильности раздражаемой ткани. Значение частоты и силы раздражения было показано на нервно-мышечном препарате. Повышение частоты и силы раздражения до известного предела вызывает увеличение высоты тетанического сокращения скелетной мышцы. Наиболее благоприятная частота нервных импульсов, поступающих в скелетную мышцу, вызывает наибольшую высоту тетануса. Эта частота называется оптимальной, или оптимумом частоты. Оптимуму частоты соответствует такая частота, при которой каждое последующее раздражение застает скелетную мышцу в состоянии наибольшей возбудимости, наблюдающейся в экзальтационной фазе. Поэтому высота каждого одиночного сокращения возрастает. Наоборот, если каждое последующее раздражение застает скелетную мышцу в фазе абсолютной рефрактерности, то тетаническое сокращение мышцы резко уменьшается или не наступает. Эта чрезмерно большая частота — наихудшая, пессимальная, или пессимум частоты. №4 Сила и работа скелетной мышцы: факторы, определяющие эти действия Сила, развиваемая мышцей или пучком мышечных волокон, является суммой сил отдельных волокон. Чем толще мышца и больше ее «физиологическая S»поперечного сечения (физиологическая сумма S поперечного сечения отдельных волокон), тем сильнее мышца. Так, например, при мышечной гипертрофии сила и толщина мышцы возрастают в одинаковой степени. Мышечная сила зависит не только от активирующего влияния со стороны ЦНС, но также определяется внешними механическими условиями, при которых работает мышца. 1.Итак, сила мышцы оценивается весом груза, который она при максимальном возбуждении способна удерживать не меняя своей длины. Сила мышцы зависит от: 1суммы сил мышечных волокон ( их сократительной способности) 2количества мышечных волокон в мышце и количества функциональных единиц, одновременно возбуждающихся при развитии напряжения 3исходной длины мышцы (предварительно растянутая мышца развивает большую силу) 4характера регулятивных явлений 5условий взаимодействия мышцы с костями скелета №5 Современные представления о механизмах мышечного сокращения и расслабления Сокращение: Этап 1. АТФазный участок головки миозина гидролизует АТФ, и головка переходит в активированное состояние (вверху справа). Однако если концентрация Са2+ в цитоплазме низка, то поперечный мостик не возникает - тропомиозин закрывает активный центр актина. Этап 2. Соединение Са2+ с тропонином С приводит к открыванию активного центра, и тогда возникает поперечный мостик (активный комплекс, внизу справа). Этап 3. При отсоединении от головки миозина АДФ осуществляется "гребковое движение" - головка сгибается, нить актина скользит относительно нити миозина, и происходит сокращение. Именно на этом этапе энергия, запасенная при расщеплении АТФ, преобразуется в механическую энергию. При этом образуется низкоэнергетический, так называемый ригорный комплекс (rigor mortis - трупное окоченение) (внизу слева). Этап 4. Присоединение к головке миозина АТФ ведет к распаду мостика, цикл завершается. Далее он повторяется, пока Са2+ связан с тропонином С. Когда же Са2+ удаляется обратно в саркоплазматический ретикулум, мышца расслабляется, а головки миозина переходят в активированное состояние. Ф - фосфат Расслабление мышцы происходит после прекращения поступления длительного нервного импульса. При этом проницаемость стенки цистерн саркоплазматической сети уменьшается, и ионы кальция под действием кальциевого насоса, используя энергию АТФ, уходят в цистерны. Удаление ионов кальция в цистерны ретикулума после прекращения двигательного импульса требует значительных энерготрат. Так как удаление ионов кальция происходит в сторону более высокой концетрации, т.е. против осмотического градиента, то на удаление каждого иона кальция затрачивается две молекулы АТФ. Концентрация ионов кальция в саркоплазме быстро снижается до исходного уровня. Белки вновь приобретают конформацию характерную для состояния покоя. Таким образом, и процесс мышечного сокращения и процесс мышечного расслабления – это активные процессы, идущие с затратами энергии в виде молекул АТФ, В гладких мышцах нет миофибрилл, которые состоят из нескольких сотен саркомеров. Тонкие нити присоединяются к сарколемме, толстые находятся внутри волокон. Ионы кальция также играют роль в сокращении, но поступают в мышцу не из цистерн, а из внеклеточного вещества, поскольку в гладких мышцах отсутствуют цистерны с ионами калькия. Этот процесс медленный и поэтому медленно работают гладкие мышцы. №6 Утомление: физиологические механизмы и принципы восстановления Утомление - это функциональное состояние, временно возникающее под влиянием продолжительной и интенсивной работы и приводящее к снижению ее эффективности. Утомление проявляется в том, что уменьшается сила и выносливость мышц, ухудшается координация движений, возрастают затраты энергии при выполнении работы одинакового характера, замедляется скорость переработки информации, ухудшается память, затрудняется процесс сосредоточения и переключения внимания, усвоения теоретического материала. Утомление связано с ощущением усталости, и в то же время оно служит естественным сигналом возможного истощения организма и предохранительным биологическим механизмом, защищающим его от перенапряжения. Утомление, возникающее в процессе упражнения, это еще и стимулятор, мобилизующий как резервы организма, его органов и систем, так и восстановительные процессы. Утомление наступает при физической и умственной деятельности. Оно может быть острым, т.е. проявляться в короткий промежуток времени, и хроническим, т.е. носить длительный характер (вплоть до нескольких месяцев); общим, т.е. характеризующим изменение функций организма в целом, и локальным, затрагивающим какую-либо ограниченную группу мышц, орган, анализатор. Различают две фазы утомления: компенсированную (когда нет явно выраженного снижения работоспособности из-за того, что включаются резервные возможности организма); некомпенсированную (когда резервные мощности организма исчерпаны и работоспособность явно снижается). Систематическое выполнение работы на фоне недовосстановления, непродуманная организация труда, чрезмерное нервно-психическое и физическое напряжение могут привести к переутомлению, а следовательно, к перенапряжению нервной системы, обострениям сердечно-сосудистых заболеваний, гипертонической и язвенным болезням, снижению защитных свойств организма. Устранить утомление возможно, повысив уровень общей и специализированной тренированности организма, оптимизировав его физическую, умственную и эмоциональную активность. Профилактике и отдалению умственного утомления способствует мобилизация тех сторон психической активности и двигательной деятельности, которые связаны с теми, что привели к утомлению. Необходимо активно отдыхать, переключаться на другие виды деятельности, использовать арсенал средств восстановления. Восстановление - процесс, происходящий в организме после прекращения работы и заключающийся в постепенном переходе физиологических и биохимических функций к исходному состоянию. Время, в течение которого происходит восстановление физиологического статуса после выполнения определенной работы, называют восстановительным периодом. Следует помнить, что в организме как во время работы, так и в предрабочем и послерабочем покое, на всех уровнях его жизнедеятельности непрерывно происходят взаимосвязанные процессы расхода и восстановления функциональных, структурных и регуляторных резервов. Во время работы процессы диссимиляции преобладают над ассимиляцией и тем больше, чем значительнее интенсивность работы и меньше готовность организма к ее выполнению. В восстановительном периоде преобладают процессы ассимиляции, а восстановление энергетических ресурсов происходит с превышением исходного уровня (сверхвосстановление, или суперкомпенсация). Это имеет огромное значение для повышения тренированности организма и его физиологических систем, обеспечивающих повышение работоспособности. Различают раннюю и позднюю фазу восстановления. Ранняя фаза заканчивается через несколько минут после легкой работы, после тяжелой - через несколько часов. Поздние фазы восстановления могут длиться до нескольких суток. Утомление сопровождается фазой понижения работоспособности, а спустя какое-то время может смениться фазой повышенной работоспособности. Длительность этих фаз зависит от степени тренированности организма, а также от выполняемой работы. Рационально сочетать нагрузки и отдых необходимо для того, чтобы сохранить и развить активность восстановительных процессов. Дополнительными средствами восстановления могут быть факторы гигиены, питания, массаж, биологически активные вещества (витамины). Главный критерий положительной динамики восстановительных процессов - готовность к повторной деятельности, а наиболее объективным показателем восстановления работоспособности служит максимальный объем повторной работы. С особой тщательностью необходимо учитывать нюансы восстановительных процессов при организации занятий физическими упражнениями и планировании тренировочных нагрузок. Повторные нагрузки целесообразно выполнять в фазе повышенной работоспособности. Слишком длинные интервалы отдыха снижают эффективность тренировочного процесса. Чтобы ускорить процесс восстановления, в спортивной практике используется активный отдых, т.е. переключение на другой вид деятельности. №7 Функциональные особенности гладких мышц •Гладкие мышцы находятся в стенках внутренних органов и кровеносных сосудов. Регуляция их тонуса и сократительной активности осуществляется эффективными волокнами симпатической и парасимпатической НС, а также местными гуморальными и физическими воздействиями. •Сократительный аппарат гладких мышц, как и скелетных, состоит из толстых миозиновых и тонких актиновых нитей. Вследствии их нерегулярного распределения клетки гладких мышц не имеют характерной для скелетной и сердечной мышцы поперечной исчерченности. •Гладкомышечные клетки имеют веретенообразную форму, l = 500-400 мкм. и толщину 2-10 мкм. Отделены друг от друга узкими щелями (60-150 нм). •Возбуждение электротонически распространяется по мышце от клетки к клетке через особые контакты (нексусы) между плазматическими мембранами соседних клеток. •Гладкая мышца с морфологической точки зрения является не истинным, а функциональным синцитием. Клетки в гладкой мышце расположены хаотично, неравномерно, иногда они собраны в пучки или тяжы. Их окружает соединительная ткань. Волокна гладких мышц •сокращаются в результате относительного скольжения миозиновых и атктиновых нитей, но скорость их сокращения и скорость расщепления АТФ в 100-1000 раз меньше, чем в скелетных мышцах. •Гладкие мышцы хорошо приспособлены к длительному тоническому сокращению без развития утомления. •При этом их энерготраты крайне невелики. По функциональным особенностям гладкие мышцы подразделяются –на мышцы, обладающие и –не обладающие спонтанной активностью. Гладкие мышцы, обладающие спонтанной активностью •способны сокращаться и при отсутствии прямых возбуждающих нервных и гуморальных воздействий( например, ритмические сокращения гладких мышц кишечника). •ПП таких клеток постоянно спонтанно колеблются в пределах 30-70 мВ («дрейф»). При снижении потенциала покоя до критического уровня возникает потенциал действия, вызывающий сокращение мышечного волокна. •Длительность ПД гладкомышечных волокон намного больше, чем у волокон поперечно-полосатой мускулатуры (несколько сек.) •Продолжительность самого сокращения также превышает несколько секунд. •Особенно медленно протекает расслабление. •Возбуждение распространяется со скоростью 2-10 см/сек, т.е. меньшей, чем по скелетной мышце. Спонтанная активность гладкомышечных клеток связана с их растяжением, вызывающим деполяризацию мембраны мышечных клеток серии импульсов с последующим сокращением. Гладкие мышцы, не обладающие спонтанной активностью сокращаются под влиянием импульсов вегетативной НС. Так, в отличие от мышц кишечника, мышечные клетки артерий, семенных протоков и радужки обладают слабой спонтанной активностью, или вообще не проявляют ее. •Отдельные нервные импульсы не способны вызвать пороговую деполяризацию таких клеток и их сокращения. ПД волокна с последующим сокращением возникает лишь при поступлении к нему серии импульсов с частотой 1 импульс в секунду и выше. •Возбуждение передается от одной клетки к последующим через плотные контакты их мембран №8 Структурно-функциональные особенности нервного волокна Нервные волокна - отростки нервных клеток (нейронов), имеющие оболочку и способные проводить нервный импульс. Главной составной частью нервного волокна является отросток нейрона, образующий как бы ось волокна. Большей частью это аксон. Нервный отросток окружен оболочкой сложного строения, вместе с которой он и образует волокно. Толщина нервного волокна в организме человека, как правило, не превышает 30 микрометров. Нервные волокна делятся на мякотные (миелиновые) и безмякотные (безмиелиновые). Первые имеют миелиновую оболочку, покрывающую аксон, вторые лишены миелиновой оболочки. Как в периферической так и в центральной нервной системе преобладают миелиновые волокна. Нервные волокна, лишенные миелина располагаются преимущественно в симпатическом отделе вегетативной нервной системы. В месте отхождения нервного волокна от клетки и в области перехода его в конечные разветвления нервные волокна могут быть лишены всяких оболочек, и тогда они называются голыми осевыми цилиндрами. В зависимости от характера проводимого по ним сигнала, нервные волокна подразделяют на двигательные вегетативные, чувствительные и двигательные соматические. |