ответы к экзамену по гистологии. Ответы к экзамену по цитологии и гистологии История создания и современное состояние клеточной теории
 Скачать 9.54 Mb. Скачать 9.54 Mb.
|

|
Мейоз (редукционное деление) - деление эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа: - редукционный, - эквационный. С уменьшением числа хромосом в результате мейоза в жизненном цикле происходит переход от диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса. В связи с тем, что в профазе первого, редукционного, этапа происходит попарное слияние (конъюгация) гомологичных хромосом, правильное протекание мейоза возможно только в диплоидных клетках или в чётных полиплоидах (тетра-, гексаплоидных и тому подобных клетках). Мейоз может происходить и в нечётных полиплоидах (три-, пентаплоидных и тому подобных клетках), но в них, из-за невозможности обеспечить попарное слияние хромосом в профазе I, расхождение хромосом происходит с нарушениями, которые ставят под угрозу жизнеспособность клетки или развивающегося из неё многоклеточного гаплоидного организма. Этот же механизм лежит в основе стерильности межвидовых гибридов. Поскольку у межвидовых гибридов в ядре клеток сочетаются хромосомы родителей, относящихся к различным видам, хромосомы обычно не могут вступить в конъюгацию. Это приводит к нарушениям в расхождении хромосом при мейозе и, в конечном счете, к нежизнеспособности половых клеток, или гамет. Определенные ограничения на конъюгацию хромосом накладывают и хромосомные мутации (масштабные делеции, дупликации, инверсии или транслокации). Фазы мейоза: Мейоз состоит из двух последовательных делений с короткой интерфазой между ними. Профаза I - профаза первого деления очень сложная и состоит из 5 стадий: 1) Фаза лептотены или лептонемы - конденсация ДНК с образованием хромосом в виде тонких нитей. 2) Зиготена или зигонема - коньюгация (соединение) гомологичных хромосом с образованием структур, состоящих из двух соединенных хромосом, называемых тетрадами или бивалентами. 3) Пахитена или пахинема - кроссинговер (перекрест) обмен участками между гомологичными хромосомами; гомологичные хромосомы остаются соединенными между собой. 4) Диплотена или диплонема - происходит частичная деконденсация хромомсом, при этом часть геном может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединенными между собой. 5) Диакинез - ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; гомологичные хромосомы остаются соединенными между собой. Метафаза I - бивалентные хромосомы выстраиваются вдоль экватора клетки. Анафаза I - микротрубочки сокращаются, биваленты делятся и хромосомы расходятся к полюсам. Важно отметить, что, из-за коньюгации хромосом в зиготене, к полюсам расходятся целые хромосомы, состоящие из двух хроматид каждая, а не отдельные хроматиды, как в митозе. Телофаза I - хромосомы деспирализуются и появляется ядерная оболочка. Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит редупликации ДНК. Профаза II - происходит конденсация хромосом, клеточный центр делится и продукты его деления расходятся к полюсам ядра, разрушается ядерная оболочка, образуется веретено деления. Метафаза II - унивалентные хромосомы (состоящие из двух хроматид каждая) располагаются на «экваторе» (на равном расстоянии от «полюсов» ядра) в одной плоскости, образуя так называемую метафазную пластинку. Анафаза II - униваленты делятся и хроматиды расходятся к полюсам. Телофаза II - хромосомы деспирализуются и появляется ядерная оболочка.В результате из одной диплоидной клетки образуется четыре гаплоидных клетки. В тех случаях, когда мейоз сопряжен с гаметогенезом (например, у многоклеточных животных), при развитии яйцеклеток первое и второе деления мейоза резко неравномерны. В результате формируется одна гаплоидная яйцеклетка и два так называемых редукционных тельца (абортивные дериваты первого и второго делений). 42. Лимфоциты и моноциты позвоночных. Их роль в воспалительных процессах Лимфоциты - это группа клеток, которые относятся к одной из разновидностей лейкоцитов. Изображения лимфоцитов: Рисунок, полученный с помощью сканирующего электронного микроскопа, обычной циркулирующей крови человека, содержащей красные клетки крови, несколько видов белых клеток крови, а именно: лимфоциты, моноциты и нейтрофил и много тромбоцитов в форме мелких дисков: Виды лимфоцитов: 1) В-лимфоциты распознают чужеродные структуры (антигены) вырабатывая при этом специфические антитела (белковые молекулы, направленные против чужеродных структур). 2) Т-лимфоциты выполняют функцию регуляции иммунитета. Т-помошники стимулируют выработку антител, а Т-супрессоры тормозят ее. 3) К-лимфоциты способны разрушать чужеродные структуры, помеченные антителами. Под влиянием этих клеток могут быть разрушены различные бактерии, раковые клетки или клетки инфицированные вирусами. 4) NK-лимфоциты осуществляют контроль над качеством клеток организма. При этом NK-лимфоциты способны разрушать клетки, которые по своим свойствам отличаются от нормальных клеток, например, раковые клетки. В норме лимфоциты составляют от 19% до 37% общего количества лейкоцитов крови. Моноциты - крупные одноядерные лейкоциты, относящиеся к агранулоцитам, то есть не содержащие в цитоплазме азурофильных гранул. Моноциты, как и лимфоциты, имеют несегментированное ядро. Но в отличие от лимфоцитов, моноциты имеют ядро неправильной формы, в то время как лимфоциты имеют круглое тёмное ядро. Моноциты это самые большие клетки крови. Попадая в ткани, они превращаются в макрофагов. Макрофаги это большие клетки, активно разрушающие бактерии. Макрофаги в больших количествах накапливаются в очагах воспаления. По сравнению с нейтрофилами моноциты более активны в отношении вирусов, чем бактерий, и не разрушаются во время реакции с чужеродным антигеном, поэтому в очагах воспаления вызванного вирусами гной не формируется. Также моноциты накапливаются в очагах хронического воспаления. Моноциты способны к активному амебоидному движению, к экстравазации (эмиграции за пределы кровеносных сосудов) и к хемотаксису (преимущественной миграции в места воспаления или повреждения тканей). Моноциты обладают выраженной способностью к фагоцитозу, причём являются макрофагами, то есть способны к поглощению достаточно крупных чужеродных частиц и клеток или большого числа мелких частиц. При этом моноциты обычно не погибают после фагоцитирования чужеродных частиц и клеток (если только чужеродная клетка или частица не имеет каких-либо цитотоксических для моноцита свойств). Этим они отличаются от микрофагов - нейтрофилов и эозинофилов, способных поглощать лишь относительно небольшие частицы и как правило погибающих после фагоцитирования. Моноциты способны, эмигрировав в ткани, превращаться в тканях в резидентные тканевые макрофаги. Моноциты также способны, подобно другим макрофагам, выполнять процессинг антигенов и представлять антигены Т-лимфоцитам для распознавания и обучения, то есть являются антигенпрезентирующими клетками иммунной системы. Моноциты секретируют растворимые цитокины, оказывающие воздействие на функционирование других звеньев иммунной системы. Цитокины, секретируемые моноцитами, называют монокинами. В норме моноциты составляют от 3% до 11% общего количества лейкоцитов крови. Относительное увеличение процента моноцитов в лейкоцитарной формуле называется относительным моноцитозом. Абсолютное увеличение числа моноцитов называется абсолютным моноцитозом. Относительное уменьшение процента моноцитов называется относительной монопенией, а абсолютное уменьшение их числа - абсолютной монопенией. Рост и созревание моноцитарно-макрофагального ростка костного мозга усиливается ГМ-КСФ и М-КСФ, тормозится глюкокортикоидами. При стрессе, шоке, терапии экзогенными глюкокортикоидами отмечается абсолютная или относительная монопения. 43. Митоз, стадии митоза Митоз (реже кариокинез или непрямое деление) - деление ядра эукариотической клетки с сохранением числа хромосом. В отличие от мейоза, митотическое деление протекает без осложнений в клетках любой плоидности, поскольку не включает как необходимый этап конъюгацию гомологичных хромосом в профазе. Фазы митоза Митоз - лишь одна из частей клеточного цикла, но он достаточно сложен, чтобы в его составе, в свою очередь, были выделены четыре фазы: 1) профаза, 2) метафаза, 3) анафаза 4) телофаза. Удвоение хромосом происходит еще в ходе интерфазы. В результате этого, в митоз хромосомы вступают уже удвоенными, напоминающими букву X (идентичные копии материнской хромосомы соединены друг с другом в области центромеры (участка хромосомы, характеризующегося специфической последовательностью нуклеотидов и структурой.)). В профазе происходит конденсация хромосом, клеточный центр делится и продукты его деления расходятся к полюсам ядра, разрушается ядерная оболочка, образуется веретено деления. В метафазе хромосомы располагаются на «экваторе» (на равном расстоянии от «полюсов» ядра) в одной плоскости, образуя так называемую метафазную пластинку. Важно отметить, что они остаются в таком положении в течение довольно длительного времени. Обычно в связи с этим метафаза - наиболее удобное время для подсчета хромосомных чисел. В анафазе хромосомы делятся (соединение в районе центромеры разрушается) и расходятся к полюсам деления. В телофазе происходит разрушение веретена деления и образование ядерной оболочки вокруг дочерних ядер и не только ядер. Ранняя анафаза митоза в клетке почки тритона (световой микроскоп, иммунофлюоресценция): Описание рисунка: микротрубочки зелёные, хромосомы голубые. Митоз: 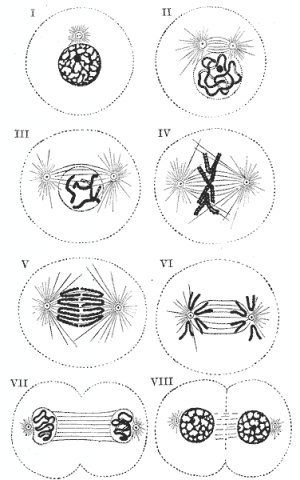 Описание рисунка: I - III - профаза; IV - метафаза; V - VI - анафаза; VII - VIII - телофаза. Варианты митоза: Следует отметить, что в разных группах живых организмов митоз протекает несколько по-разному. Описанный выше вариант митоза называется открытым ортомитозом (ядерная оболочка разрушается, веретено деления прямое, поскольку продукты деления клеточного центра располагаюся на противоположных полюсах ядра). Характерен для многоклеточных животных, многоклеточных растений и ряда простейших. В некоторых группах простейших продукты деления клеточного центра в анафазе не достигают противоположных сторон ядра, в результате чего микротрубочки веретена деления располагаются под углом, напоминая букву V (такой вариант деления получил название плевромитоз). В ряде случаев митоз происходит без разрушения ядерной оболочки (закрытый митоз). Наконец, клеточный центр может содержать центриоли (как, например, у животных) или не содержать их (как, например, у цветковых растений). Соответственно, различают также центриолярный и ацентриолярный митоз. Амитоз (прямое деление) - деление клетки без образования веретена деления. Долгое время наряду с митозом и мейозом считался одним из основных способов деления клеток. Это понятие ещё фигурировало в некоторых учебниках до 1980-х гг. В настоящее время считается, что все явления, относимые к амитозу - результат неверной интерпретации недостаточно качественно приготовленных микроскопических препаратов, или интерпретации как деления клетки явлений, сопровождающих разрушение клеток или иные патологические процессы. 44. Строение костной ткани Костная ткань - одна из разновидностей соединительной ткани. Отличается высокой минерализацией межклеточного вещества. Минеральные структуры формируется на белке коллагене, трехчастная спиральная структура которого является матрицей для отложения минеральных форм кости. Кость служит основным депо кальция в организме и активно участвует в кальциевом обмене.Снаружи лежит надкостница. Она состоит из плотной волокнистой соединительной ткани и молодых костных клеток - остеобластов. В надкостнице много кровеносных сосудов. Остеобласты, выделяя костную ткань и замуровываясь в ней, превращаются в остеоциты. Под надкостницей в несколько рядов располагаются параллельные костные пластинки. Такие же, но внутренние костные пластинки лежат со стороны костного мозга. Остеоциты лежат между пластинками. Они имеют много отростков, проникающих в костные канальцы. В среднем слое костной пластинки видны концентрические костные пластинки, окружающие канал, где проходит кровеносный сосуд. Это образование называется остеоном или гаверсовой системой. Между остеонами располагаются вставочные пластинки. 45. Клетки прокариот и эукариот. Различия и сходство их строения. Все организмы, имеющие клеточное строение, делятся на две группы: предъядерные (прокариоты) и ядерные (эукариоты). Клетки прокариот, к которым относятся бактерии, в отличие от эукариот, имеют относительно простое строение. В про-кариотической клетке нет организованного ядра, в ней содержится только одна хромосома, которая не отделена от остальной части клетки мембраной, а лежит непосредственно в цитоплазме. Однако в ней также записана вся наследственная информация бактериальной клетки. Цитоплазма прокариот по сравнению с цитоплазмой эука-риотических клеток значительно беднее по составу структур. Там находятся многочисленные более мелкие, чем в клетках эукариот, рибосомы. Функциональную роль митохондрий и хло-ропластов в клетках прокариот выполняют специальные, довольно просто организованные мембранные складки. Клетки прокариот, так же как и эукариотические клетки, покрыты плазматической мембраной, поверх которой располагается клеточная оболочка или слизистая капсула. Несмотря на относительную простоту, прокариоты являются типичными независимыми клетками. Сравнительная характеристика клеток эукариот. По строению различные эукариотические клетки сходны. Но наряду со сходством между клетками организмов различных царств живой природы имеются заметные отличия. Они касаются как структурных, так и биохимических особенностей. Для растительной клетки характерно наличие различных пластид, крупной центральной вакуоли, которая иногда отодвигает ядро к периферии, а также расположенной снаружи плазматической мембраны клеточной стенки, состоящей из целлюлозы. В клетках высших растений в клеточном центре отсутствует центриоль, встречающаяся только у водорослей. Резервным питательным углеводом в клетках растений является крахмал. В клетках представителей царства грибов клеточная стенка обычно состоит из хитина - вещества, из которого построен наружный скелет членистоногих животных. Имеется центральная вакуоль, отсутствуют пластиды. Только у некоторых грибов в клеточном центре встречается центриоль. Запасным углеводом в клетках грибов является гликоген. В клетках животных отсутствует плотная клеточная стенка, нет пластид. Нет в животной клетке и центральной вакуоли. Центриоль характерна для клеточного центра животных клеток. Резервным углеводом в клетках животных также является гликоген. 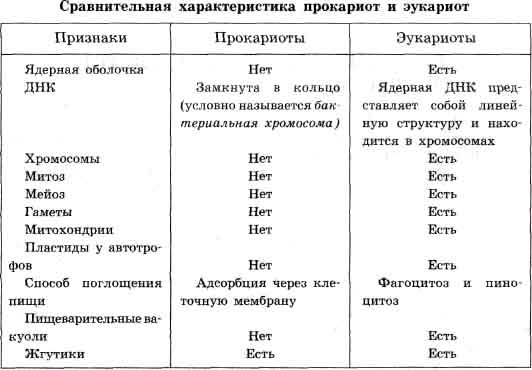 46. Строение и функции эритроцитов позвоночных. Эритроциты - красные кровяные тельца - клетки крови человека, позвоночных животных и некоторых беспозвоночных (иглокожих). Основной функцией эритроцитов является перенос кислорода из лёгких к тканям тела, и транспорт диоксида углерода (углекислого газа) в обратном направлении. Однако, кроме участия в процессе дыхания, они выполняют в организме также следующие функции: - участвуют в регулировке кислотно-щелочного равновесия; - поддерживают изотонию крови и тканей; - адсорбируют из плазмы крови аминокислоты, липиды и переносят их к тканям. Формирование эритроцитов (эритропоэз) происходит в костном мозге черепа, ребер и позвоночника, а у детей - еще и в костном мозге в окончаниях длинных костей рук и ног. Прежде чем выйти в кровь, эртироциты последовательно проходят несколько стадий пролиферации и дифференцировки в составе эритрона - красного ростка кроветворения. а) Из стволоволовых гемопоэтических клеток сначала появляется большая клетка с ядром, не обладающая характерным красным цветом - мегалобласт. б) Затем она окрашивается в красный цвет - теперь это эритробласт. в) Уменьшается в размере в процессе развития - теперь это нормоцит. г) Утрачивает ядро - теперь это ретикулоцит. У птиц, пресмыкающихся, земноводных и рыб ядро просто теряет активность, но сохраняет способность реактивации. Одновременно с исчезновением ядра по мере взросления эритроцита из его цитоплазмы исчезают рибосомы и другие компоненты, участвующие в синтезе белка. Ретикулоциты попадают в кровеносную систему и через несколько часов становятся полноценными эритроцитами. Структура и состав Обычно эритроциты имеют форму двояковогнутого диска и содержат в основном дыхательный пигмент гемоглобин. У некоторых животных (например, верблюда) эритроциты имеют овальную форму. Содержимое эритроцита представлено главным образом дыхательным пигментом гемоглобином, обусловливающим красный цвет крови. Однако на ранних стадиях количество гемоглобина в них мало, и на стадии эритробластов цвет клетки синий; позже клетка становится серой и, лишь полностью созрев, приобретает красную окраску. Эритроциты (красные кровяные тельца крови) человека.Важную роль в эритроците выполняет клеточная (плазматическая) мембрана, пропускающая газы, ионы и воду. На поверхности липопротеидной мембраны находятся специфические антигены гликопротеидной природы - агглютиногены - факторы систем групп крови (на данный момент изучено более 15 систем групп крови: AB0, резус фактор, Даффи, Келл, Кидд), обусловливающие агглютинацию эритроцитов. Эффективность функционирования гемоглобина зависит от величины поверхности соприкосновения эритроцита со средой. Суммарная поверхность всех эритроцитов крови в организме тем больше, чем меньше их размеры. У низших позвоночных эритроциты крупные (например, у хвостатого земноводного амфиумы - 70 мкм в диаметре), эритроциты высших позвоночных мельче (например, у козы - 4 мкм в диаметре). У человека диаметр эритроцита составляет 7,2 - 7,5 мкм, толщина - 2 мкм, объём - 88 мкм3. |