ответы к экзамену по гистологии. Ответы к экзамену по цитологии и гистологии История создания и современное состояние клеточной теории
 Скачать 9.54 Mb. Скачать 9.54 Mb.
|

|
Часть рецепторов: световые, звуковые, обонятельные, вкусовые, тактильные, температурные, воспринимающие раздражения от внешней среды, - расположена вблизи внешней поверхности тела. Их называют экстерорецепторами. Другие же рецепторы воспринимают раздражения, связанные с изменением состояния и деятельности органов я внутренней среды организма. Их называют интерорецепторами (к числу интерорецепторов относят рецепторы, находящиеся в скелетной мускулатуре, их называют проприорецепторами). Эффекторные нейроны по своим идущим на периферию отросткам - афферентным, или центробежным, волокнам - передают импульсы, изменяющие состояние и деятельность различных органов. Часть эффекторных нейронов расположена в центральной нервной системе - в головном и спинном мозгу, и на периферию идет от каждого нейрона только один отросток. Таковы моторные нейроны, вызывающие сокращения скелетной мускулатуры. Часть же эффекторных нейронов целиком расположена на периферии: они получают импульсы из центральной нервной системы и передают их к органам. Таковы образующие нервные ганглии нейроны вегетативной нервной системы. Контактные нейроны, расположенные в центральной нервной системе, выполняют функцию связи между различными нейронами. Они служат как бы релейными станциями, производящими переключение нервных импульсов с одних нейронов на другие. Взаимосвязь нейронов составляет основу для осуществления рефлекторных реакций. При каждом рефлексе нервные импульсы, возникшие в рецепторе при его раздражении, передаются по нервным проводникам в центральную нервную систему. Здесь или непосредственно, или же через посредство контактных нейронов нервные импульсы переключаются с рецепторного нейрона на эффекторный, от которого они идут на периферию к клеткам. Под влиянием этих импульсов клетки изменяют свою деятельность. Импульсы, поступающие в центральную нервную систему с периферии или же передаваемые от одного нейрона другому, могут вызывать не только процесс возбуждения, но и противоположный ему процесс - торможение. Тигроидная субстанция Нисля - это специфическая структура нервных клеток выявляется при окраске основными красителями в виде базофильных глыбок, поэтому окрашенные нервные клетки имеют пятнистый вид. Она представляет собой гранулярную эндоплазматическую сеть с множеством рибосом. В этих участках идет активный синтез белка. Морфология глыбок меняется в зависимости от функционального состояния нейрона. Тигроидная субстанция Нисля: 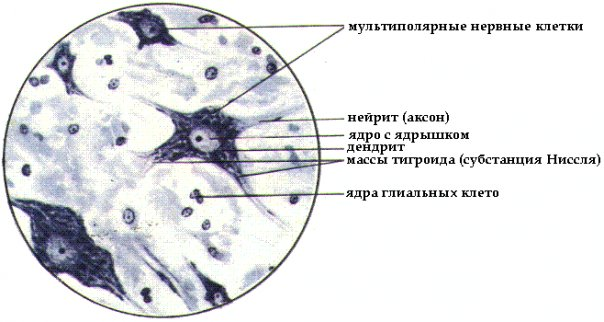 29. Строение и функции ядерной оболочки Ядерная оболочка - мембранный барьер, отделяющий ядро от цитоплазмы. Ядерная оболочка характерна для всех эукариотических клеток. Ядерная оболочка состоит из внешней и внутренней мембран, разделенных перинуклеарным пространством шириной от 20 до 60 нм. В состав ядерной оболочки входят мембрана, ядерный поровый комплекс и ламина. Ядерная оболочка образована внешней и внутренней мембранами. Наружная мембрана переходит в шероховатый эндоплазматический ретикулум, и обеспечивает присоединение структурных элементов цитоплазмы. Внутренняя выстлана белками – ламининами, образующими ядерную пластинку, которая закрепляет различные ядерные структуры. Между мембранами располагается перинуклеарное пространство. В состав ядерной оболочки также входит ядерный поровый комплекс - восмиугольный цилиндр. Ламина представляет собой густую сеть выстилающую внутреннюю мембрану изнутри и состоящую из промежуточных филаментов - ламинов и взаимодействующих с ними белков. Ламина необходима для сохранения целостности ядра. Ламина – структура и образованная белками ламинами А, B, C. A и C – производные одного гена образующиеся при альтернативном спайсинге. Ламин А содержит 113 добавочных аминокислот. Ламин В связан с липидным слоем изопрениловым якорем. Три ламины образуют димеры, имеющие стержневидный домен из а-спиралей и глобулярные домены на каждом конце. Эти домены полимеризуются и образуют сетеподобную структуру. Функции ядерной оболочки: Ядерная оболочка контролирует перемещение макромолекул между нуклеоплазмой и цитозолем, участвует в заякоревании хромосом и цитоскелета, являясь частью регуляторного механизма экспрессии у эукариот. Мутации в белках ядерной оболочки проявляются в виде различных заболеваний, таких как мышечная дистрофия, нейропатия, липидодистрофия, преждевременное старение. 30. Строение гладкой мышечной ткани. По происхождению различают три группы гладких (неисчерченных) мышечных тканей: 1) Гладкая мышечная ткань мезенхимного происхождения Развивается этот тип ткани из мезенхимы и имеет клеточное строение. Волокна мезенхимной мышечной ткани состоят из клеткок - миоцитов веретенообразной формы. Ядро расположено в центре клетки, имеет эллипсоидную форму. Соединительнотканные прослойки из коллагеновых и эластических волокон разбивают мышечную ткань на пучки первого и второго порядков. Схема строения гладкой мышечной ткани: 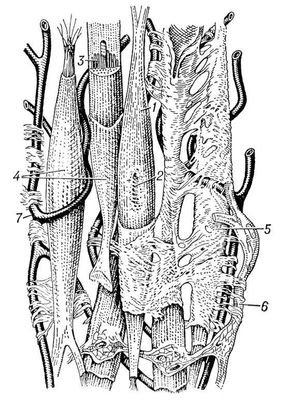 Описание рисунка: 1 - гладкая мышечная клетка; 2 - её ядро; 3 - миофибриллы; 4 - сарколемма; 5 - соединительная ткань; 6 - нерв; 7 - кровеносный капилляр. 2) Гладкая мышечная ткань эпидермального происхождения Миоэпителиальные клетки развиваются из эпидермального зачатка. Они встречаются в потовых, молочных, слюнных и слезных железах и имеют общих предшественников с железистыми секреторными клетками. Большинство миоэпителиальных клеток имеют звездчатую форму. Эти клетки нередко называют корзинчатыми: их отростки охватывают концевые отделы и мелкие протоки желез. В теле клетки располагаются ядро и органеллы общего значения, а в отростках - сократительный аппарат, организованный, как и в клетках мышечной ткани мезенхимного типа 3) Гладкая мышечная ткань нейрального происхождения Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы - суживающую и расширяющую зрачок. 31. Хроматин. Его химическая структура и уровни спирализации в клетке Хроматин - молекулы хромосомной ДНК в ядре эукариотической клетки в комплексе со специфическими белками, необходимыми для хранения, репликации и реализации генетического материала. Основную массу составляют "белки хранения", так называемые гистоны. Из этих белков построены нуклеосомы, структуры на которые намотаны нити молекул ДНК. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из белков четырех типов: H2A, H2B, H3 и H4. В одну нуклеосому входят по два белка каждого типа - всего восемь белков. Гистон H1, более крупный чем другие гистоны, связывается с ДНК в месте ее входа на нуклеосому. Нуклеосома вместе с H1 называется хроматосомой. Нить ДНК с нуклеосомами образует нерегулярную соленоидподобную структуру толщиной около 30 нанометров, так называемую 30 нм фибриллу. Дальнейшая упаковка этой фибриллы может иметь различную плотность. Если хроматин упакован плотно его называют конденсированным или гетерохроматином, он хорошо видим под микроскопом. ДНК находящаяся в гетерохроматине не транскрибируется, обычно это состояние характерно для незначащих или молчащих участков. В интерфазе гетерохроматин обычно располагается по периферии ядра (пристеночный гетерохроматин). Полная конденсация хромосом происходит перед делением клетки. Если хроматин упакован неплотно, его называют эу- или интерхроматином. Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется наличием транскрипционной активности. Плотность упаковки хроматина во многом определяется модификациями гистонов - ацетилированием и фосфориллированием. Считается, что в ядре существуют так называемые функциональные домены хроматина (ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную "территорию". К сожалению, вопрос пространственного распределения хроматина в ядре изучен пока недостаточно. Известно, что теломерные (концевые) и центромерные (отвечающие за связывание сестринских хроматид в митозе) участки хромосом закреплены на белках ядерной ламины. 32. Особенности строения мышечной ткани сердца. Атипические волокна миокарда. Мышечная ткань сердца - это поперечно-полосатая мышца. Но мышечное волокно в этой мышце составлено из клеток. Вставочные диски как бы пересекают эти волокна. Они представляют собой границы соседних клеток, то есть миофибриллы одной клетки не проникают в соседнюю. Каждая мышечная клетка имеет сарколемму, саркоплазму с располагающимися в ней миофибриллами и овальное ядро, лежащее в центре волокна. Волокна ветвятся, образуя сеть. Зарисовать участок продольного среза миокарда. Свойства сердечной мышцы: 1) Автоматия - это способность к ритмическому сокращению без всяких внешних воздействий под влиянием импульсов, возникающих в самом сердце. Ярким проявлением этого свойства сердца является способность извлеченного из организма сердца при создании необходимых условий сокращаться в течение часов и даже суток. Природа автоматии до сих пор до конца не выяснена. Но однозначно ясно, что возникновение импульсов связано с деятельностью атипических мышечных волокон, заложенных в некоторых участках миокарда. Внутри атипических мышечных клеток спонтанно генерируются электрические импульсы определенной частоты, распространяющиеся затем по всему миокарду. Первый такой участок находится в области устьев полых вен и называется синусный, или синоатриальныйузел. В атипических волокнах этого узла спонтанно возникают импульсы с частотой 60 - 80 раз в минуту. Он является главным центром автоматии сердца. Второй участок находится в толще перегородки между предсердиями и желудочками и называется предсердно-желудочковый, или атриовентрикулярный узел. Третий участок - это атипические волокна, составляющие пучок Гиса, лежащий в межжелудочковой перегородке. От пучка Гиса берут начало тонкие волокна атипической ткани - волокна Пуркинье, ветвящиеся в миокарде желудочков. Все участки атипической ткани способны генерировать импульсы, но их частота самая высокая в синусном узле, поэтому его называют водителем ритма первого порядка (пейсмекером первого порядка), и все другие центры автоматии подчиняются этому ритму. Совокупность всех уровней атипической мышечной ткани составляют проводящую систему сердца. Благодаря проводящей системе волна возбуждения, возникшая в синусном узле, последовательно распространяется по всему миокарду. 2) Возбудимость сердечной мышцы заключается в том, что под действием различных раздражителей (химических, механических, электрических и др.) сердце способно приходить в состояние возбуждения. В основе процесса возбуждения лежит появление отрицательного электрического потенциала на наружной поверхности мембран клеток, подвергшихся действию раздражителя. Как и в любой возбудимой ткани, мембрана мышечных клеток (миоцитов) поляризована. В покое она снаружи заряжена положительно, изнутри - отрицательно. Разность потенциалов определяется различной концентрацией ионов N а + и К + по обе стороны мембраны. Действие раздражителя увеличивает проницаемость мембраны для ионов К + и Nа + , происходит перестройка мембранного потенциала(калий - натриевый насос), в результате возникает потенциал действия, распространяющийся и на другие клетки. Таким образом происходит распространение возбуждения по всему сердцу. Импульсы, возникшие в синусном узле, распространяются по мускулатуре предсердий. Дойдя до атриовентрикулярного узла, волна возбуждения распространяется по пучку Гиса, а затем по волокнам Пуркинье. Благодаря проводящей системе сердца наблюдается последовательное сокращение частей сердца: сначала сокращаются предсердия, затем желудочки (начиная с верхушки сердца волна сокращения распространяется к их основанию). Особенность атриовентрикулярного узла - проведение волны возбуждения только в одном направлении: от предсердий к желудочкам. 3) Сократимость - это способность миокарда сокращаться. Оно основано на способности самих клеток миокарда отвечать на возбуждение сокращением. Это свойство сердечной мышцы определяет способность сердца выполнять механическую работу. Работа сердечной мышцы подчиняется закону "все или ничего".Суть этого закона состоит в следующем: если на сердечную мышцу наносить раздражающее действие различной силы, мышца отвечает каждый раз максимальным сокращением ("все"). Если сила раздражителя не достигает порогового значения, то сердечная мышца не отвечает сокращением ("ничего"). 33. Ядрышко. Его строение и функции Ядрышко находится внутри ядра, и не имеет собственной мембранной оболочки, однако хорошо различимо под световым и электронным микроскопом. Основной функцией ядрышка является синтез рибосом. В геноме клетки имеются специльные участки, так называемые ядрышковые организаторы, содержащие гены рибосомной РНК (рРНК), вокруг которых и формируются ядрышки. В ядрышке происходит синтез рРНК РНК полимеразой I, ее созревание, сборка рибосомных субчастиц. В ядрышке локализуются белки, принимающие участие в этих процессах. Некоторые из этих белков имеют специальную последовательность - сигнал ядрышковой локализации. Схема синтеза рибосом в клетках эукариот: 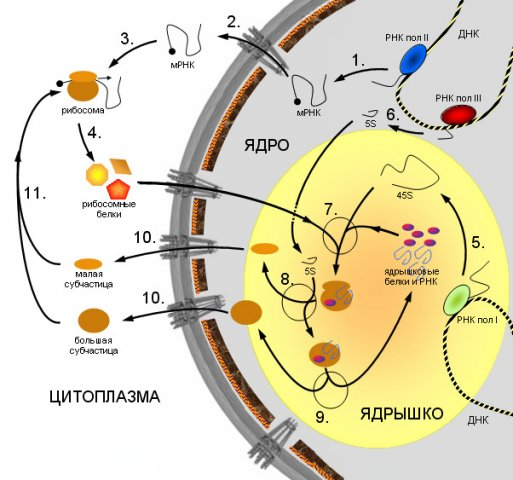 Описание рисунка: 1 - синтез мРНК рибосомных белков РНК полимеразой II; 2 - экспорт мРНК из ядра; 3 - узнавание мРНК рибосомой; 4 - синтез рибосомных белков; 5 - синтез предшественника рРНК (45S - предшественник) РНК полимеразой I; 6 - синтез 5S pРНК РНК полимеразой III; 7 - сборка большой рибонуклеопротеидной частицы, включающей 45S-предшественник, импортированные из цитоплазмы рибосомные белки, а также специальные ядрышковые белки и РНК, принимающие участие в созревании рибосомных субчастиц; 8 - присоединение 5S рРНК, нарезание предшественника и отделение малой рибосомной субчастицы; 9 - дозревание большой субчастицы, высвобождение ядрышковых белков и РНК; 10 - выход рибосомных субчастиц из ядра; 11 - вовлечение их в трансляцию. Самая высокая концентрация белка в клетке наблюдается именно в ядрышке. В этих структурах было локализовано около 600 видов различных белков, причем считается, что лишь небольшая их часть действительно необходима для осуществления ядрышковых функций, а остальные попадают туда не специфически. Под электронным микроскопом в ядрышке выделяют несколько субкомпартментов. Так называемые Фибриллярные центры окружены участками плотного фибриллярного компонента, где и происходит синтез рРНК. Снаружи от плотного фибриллярного компонента расположен гранулярный компонент, представляющий собой скопление созревающих рибосомных субчастиц. 34. Строение поперечно-полосатой мышечной ткани Поперечно-полосатая мышечная ткань происходит из мезодермы. Она имеет симпластическое строение, то есть границы между клетками отсутствуют. Ее структурная и функциональная единица - мышечное волокно, имеющее поперечную исчерченность. Волокна ограничены оболочкой - сарколеммой, под которой располагаются ядра. В цитоплазме (саркоплазме) мышечного волокна упорядоченно располагаются миофибриллы. Они имеют чередующиеся участки с разными оптическими свойствами: диски А - анизотропные (темные) и диски И – изотропные (светлые). Волокна поперечно-полосатых мышц представляют собой многоядерные гигантские клетки цилиндрической формы: их длина колеблется от нескольких милиметров до нескольких сантиметров. Их толщина составляет от 10 до 150 мкм. Сарколемма достаточно развита, вдоль нее раположены многочисленные ядра, неравномерно разбросанные в пределах волокна. Они имеют овальную форму на продольном срезе и округлую на поперечном. Волокна характеризуются как продольной, так и поперечной исчерченностью, которая выражена значительнее. Наличие миофибрилл, идущих вдоль волокна, обусловливает продольную исчерченность. Миофибриллы состоят из чередующихся светлых изотропных и темных анизотропных полос; светлые называются А-полосами, а темные - I-полосами. Эти полосы плотно уложены, что придает мышечному волокну вид поперечной исчерченности. Окончания волокон тупо закруглены. Любая поперечно-полосатая мышца, например двуглавые мышцы, одета в покров из соединительной ткани, состоящий из трех компонентов. Эпимизий из соединительной ткани, содержащей большое количество жировых клеток, покрывает мышцу (влагалище мышцы) и, погружаясь, разделяет брюшко мышцы на пучки. Каждый пучок волокон в пределах мышцы окружен толстым слоем соединительной ткани - перимизием. В пределах пучка каждое поперечно-полосатое мышечное волокно окружено тонкой прослойкой, богатой кровеносными сосудами - эндомизием. С помощью электронной микроскопии установлено, что волокна поперечно- полосатых мышц имеют сложное строение. Миофибриллы состоят из двух типов, смещающихся относительно друг друга, миофиламентов: тонких актин-содержащих и толстых миозин-содеращих. I-полосы состоят из тонких миофиламентов, а А-полосы - из обоих типов. Точно по центру каждой I-полосы расположена темная Z-линия, к каждой стороне которой прикрепляются актин-содержащие тонкие миофиламенты. Расстояние между каждой парой Z-линий называется саркомером, элементарная сократимая единица поперечно-полосатых мышц. Тонкие миофиламенты выступают в область А-полосы, причем один конец миофиламента свободен и находится между толстыми миофиламентами, что способствует возникновению светлой Н-зоны в середине А-полосы. Толстые миофиламенты тянутся через всю А-полосу и их концы свободны. Миофибриллы расположены в строго определенном порядке: каждый толстый миофиламент окружен шестью равноудаленными тонкими миофиламентами, которые расположены в форме шестиугольника. При сокращении, длина миофиламентов не уменьшается, а увеличивается только лишь их степень смещения относительно друг друга. В результате этого, I-полоса уменьшается и Н-зона тоже становится очень узкой, однако длина А-полосы практически не изменяется. На поперечном срезе миофибрилла имеет 3 вида, в зависимости от уровня среза: только на уровне тонких миофиламентов, толстых миофиламентов или на уровне двух типов филаментов одновременно. Две эллиптические митохондрии окружают каждую миофибриллу на уровне I-полосы. При исследовании под электронным микроскопом удается выяснить, что сарколемма состоит из плазматической мембраны и вместе с хорошо развитой базальной пластинкой и сетью тонких ретикулиновых волокон. Плазматическая мембрана вдавливается и проходит через волокно на уровне границы между А- и I-полосами в виде вытянутой трубочки (Т-трубочка). При сокращении волна деполяризации распространяется вдоль сарколеммы и благодаря Т-трубочкам более или менее синхронно достигает всех частей миофибриллы. Саркоплазматический гладкий эндоплазматический ретикулум опоясывает каждую миофибриллу несколько раз таким образом, что по бокам каждой Т-трубочки параллельно располагаются две терминальные цистерны. Эти структуры, видимые рядом в поперечном разрезе, называются триадой. Трубочки и цистерны обеспечивают изоляцию зоны катионов кальция, что происходит во время расслабления мышцы. Большинство мышц человека содержит три типа поперечно-полосатых волокон: красные, белые и промежуточные. Преобладающим типом являются красные волокна, которые характеризуются малой толщиной и обилием пигментированного белка - миоглобина, и относительно небольшим количеством миофибрилл. Красные, или слабо сокращающиеся волокна, выполняют повторяющиеся сократительные движения и характерны для мышц туловища, ответственных за поддержание позы. Белые волокна толще и содержат большее количество миофибрилл, но меньше миоглобина, чем красные волокна. Они способны сокращаться быстрее и сравнительно быстро устают, поэтому они больше подходят для коротких вспышек активности. Промежуточные волокна по размерам и характеристикам занимают промежуточное положение между красными и белыми волокнам. 35. Жизненный цикл клетки. Три категории клеток, различающиеся в отношении жизненного цикла Жизненный, цикл клетки - это время существования клетки от деления до следующего деления, или от деления до смерти. Для разных типов клеток клеточный цикл различен. Примерный жизненный цикл клетки: 1) Рост и стадия размножения в недифференцированном состоянии. 2) Стадия дифференцировки. 3) Дифференцировка. 4) Стадия нормальной активности. 5) Стадия старения. 6) Дезинтеграция. Естественная смерть. В организме млекопитающих и человека различают следующие три группы клеток, локализующиеся в разных тканях и органах: 1) часто делящиеся клетки (малодифференцированные клетки эпителия кишечника, базальные клетки эпидермиса и другие); 2) редко делящиеся клетки (клетки печени - гепатоциты); 3) неделящиеся клетки (нервные клетки центральной нервной системы, меланоциты и другие). Жизненный цикл у этих клеточных типов различен. Жизненный цикл у часто делящихся клеток - это время их существования от начала деления до следующего деления. Жизненный цикл таких клеток нередко называют митотическим циклом. Такой клеточный цикл подразделяется на два основных периода: 1) митоз или период деления; 2) интерфаза - промежуток жизни клетки между двумя делениями. 36. T- и B- лимфоциты их роль в реакциях гуморального и клеточного иммунитета. В организме взрослого человека 25 - 40% всех лейкоцитов крови составляют лимфоциты (1000 - 3600 клеток в 1 мкл), у детей - 50%. Состояние, при котором их число увеличивается, называется лимфоцитозом, уменьшается - лимфопенией. Лимфоциты обладают уникальным свойством - способностью распознавать антигены. Лимфоциты образуются в: - лимфатических узлах, -миндалинах, - пейеровых бляшках, - червеобразном отростке, - селезенке, - вилочковой железе (тимусе), - костном мозге. При добавлении к культуре лимфоцитов растительного белка фитогемагглютинина лимфоциты значительно увеличиваются, начинают делиться (митоз), усиленно синтезировать РНК, ДНК, белки и ферменты. Большинство покоящихся лимфоцитов представляют собой малые лимфоциты - небольшие клетки с темным ядром, что обусловлено конденсацией хроматина и сравнительно небольшим количеством цитоплазмы, содержащей разрозненные митохондрии. Эти лимфоциты участвуют в иммунном ответе и представлены двумя главными классами: 1) B-лимфоцитами (10-15% лимфоцитов крови) - функциональный тип лимфоцитов, играющих важную роль в обеспечении гуморального иммунитета. Буква "B" в названии связана с тем, что клетки были впервые обнаружены в фабрициевой сумке (bursa fabricii) птиц. У эмбрионов человека и других млекопитающих B-лимфоциты образуются в печени и костном мозге из стволовых клеток, а у взрослых млекопитающих - в красном костном мозге. При контакте с антигеном или стимуляции со стороны T-клеток некоторые B-лимфоциты трансформируются в плазматические клетки, способные к продукции антител. Другие активированные B-лимфоциты превращаются в B-клетки памяти. 2) T-лимфоцитами (70-80% лимфоцитов крови) - лимфоциты, которые у взрослых людей и млекопитающих созревают в тимусе. Играют важную роль в адаптивном то есть приобретённом иммунном ответе. Обеспечивают распознавание и уничтожение клеток, несущих чужеродные антигены, усиливают действие моноцитов, NK-клеток, а также принимают участие в переключении изотипов иммуноглобулинов то есть в изменении продукции ранних иммуноглобулинов IgM, на поздние IgG, IgE, IgA B-клетками. Существует несколько видов Т-лимфоцитов: - Т-киллеры - вид лимфоцитов, осуществляющий лизис клеток-мишеней, к которым можно отнести возбудителей инфекционных болезней, грибки, микобактерии, опухолевые клетки и прочие антигены. Функции Т-киллеров во многом сходны с В-киллерами, чьим общим предком являются так называемые "ни Т ни В - лимфоциты" (0-лимфоциты). Действие Т-киллеров усиливают Т-супрессоры. Также cуществуют "двойные" лимфоциты, несущие на себе маркеры и Т и В - киллеров. - Т-супрессоры регулируют самые различные формы гуморального и клеточного иммунного ответа, включая гиперчувствительность замедленного типа, пролиферацию цитотоксических лимфоцитов и пролиферацию антигенспецифических лимфоцитов. Однако системы супрессии далеко не просты. - Т-регуляторы (Т-дифференцирующие, Td-лимфоциты) - центральные регуляторы иммунного ответа. Основная их функция - контролировать силу и продолжительность иммунного ответа через регуляцию функции Т-эффекторных клеткок (Т-хелперов и Т-цитотоксических клеток). Эти клетки экспрессирут транскрипционный фактор, регулирующий транскрипцию генов ответственных за дифференцировку Т-клеток и экспрессию цитокинов и других факторов, участвующих в супрессии иммуного ответа. - Т-амплифайеры выделяют и усиляют соединения, которые являются стимуляторами роста для всех видов Т-лимфоцитов и активаторами К-клеток. - Т-контрсупрессоры. - Т-клетки памяти хранят информацию о ранее действовавших антигенах и формируют вторичный имунный ответ, осуществляющийся в более короткие сроки, чем первичный имунный ответ, так как минует основные стадии этого процесса. - Т-хелперы - помощники. Т-хелперы участвуют в адаптивном иммунном ответе. Существует несколько типов Т-хелперов: Т-хелперы 1-го типа - Th1, Т-хелперы 2-го типа - Th2 и Т-хелперы 17 - Th17. Оставшиеся лимфоциты называются нулевыми клетками. Кроме того, можно выделить субпопуляцию нормальных киллеров. Нормальные киллеры (NK) - это большие зернистые лимфоциты с характерной морфологией: основная часть обильной цитоплазмы содержит несколько митохондрий, свободные рибосомы с отдельными элементами шероховатого эндоплазматического ретикулума, аппарат Гольджи и характерные электроноплотные гранулы, связанные с мембраной. Большие зернистые лимфоциты с активностью нормальных киллеров выполняют цитотоксические функции, также как и цитотоксические T-лимфоциты. В образовании антител центральная роль принадлежит B-лимфоцитам. При этом B-лимфоциты обеспечивают специфический приобретенный иммунитет совместно с другими малыми лимфоцитами - T-лимфоцитами, используя разнообразные механизмы, направленные в большинстве случаев на расширение пределов эффективности врожденного иммунитета. Свои функции воздействия на другие клетки T-лимфоциты осуществляют путем выделения цитокинов, которые передают сигналы другим клеткам, или в результате прямых межклеточных контактов. Как и в случае B-лимфоцитов, отбор и активация T-лимфоцитов происходят после контакта с антигеном, затем они проходят стадию клональной экспансии и превращаются в зрелые T-хелперы и цитотоксические T-лимфоциты, а также формируют обширную популяцию клеток памяти. Одна из важных регуляторных функций T-лимфоцитов - это их способность стимулировать B-клетки к пролиферации и дифференцировке. Другая важная регуляторная функция T-клеток состоит в их способности угнетать иммунный ответ. При этом T-хелперы и T-супрессоры обнаруживают комплексный тип антигенной специфичности. Фундаментальным свойством T-клеток является их специфичность по отношению к продуктам главного комплекса гистосовместимости. 37. Клеточная патология. Теория паранекроза (Д. Н. Насонов и В. Я. Яковлев). Целлюлярная патология - клеточная патология - теория медицины, основанная на учении Р. Вирхова о клетке как материальном субстрате болезни. Сформулирована Вирховом в 1855 - 1858. Имела целью преодоление односторонности гуморальной патологии и так называемых солидарных, в том числе "невристических", концепций, видевших причины всех болезней в изменениях плотных частиц и нарушениях нервной системы. Естественнонаучной и методической основой Целлюлярная патология были клеточная теория строения организмов и микроскопическая техника. |