Иммунология. Ответы на вопросы Лимфоциты Лимфоциты
 Скачать 82.91 Kb. Скачать 82.91 Kb.
|

|
Ответы на вопросы: 1. Лимфоциты Лимфоциты – это крупные кровяные клеточные строения, принадлежащие к типу клеток лейкоцитов. Они исполняют защитную функцию: борются с патогенными вирусами, бактериями, микроорганизмами в теле пациента, оберегают организм от злокачественных опухолей. Лимфоциты в крови – это главный защитник иммунитета организма. 2.Виды лимфоцитов По функциональным признакам различают три типа лимфоцитов: B-клетки, T-клетки, NK-клетки. B-лимфоциты или B-клетки распознают чужеродные структуры (антигены), вырабатывая при этом специфические антитела (белковые молекулы, направленные против конкретных чужеродных структур). Т-лимфоциты или Т-клетки включают в себя Т-цитотоксические лимфоциты, Т-хелперы, Т-регуляторы. Т-цитотоксические лимфоциты выполняют киллерные функции, Т-хелперы регулируют и направляют иммунитет, а Т-регуляторы тормозят излишний иммунный ответ. NK-лимфоциты способны уничтожать клетки, признаваемые организмом генетически-чужеродными или потерявшие отличительные признаки "своего" (без учета антигенной специфичности). Содержание Т-лимфоцитов в крови составляет 65—80 % от общего числа лимфоцитов, В-лимфоцитов — 8—20 %, NK-лимфоцитов — 5—20 % 3. Какой иммунитет формируют? Обеспечивают гуморальный иммунитет (выработка антител), клеточный иммунитет (контактное взаимодействие с клетками-мишенями), а также регулируют деятельность клеток других типов. 4. Активация лимфоцитов 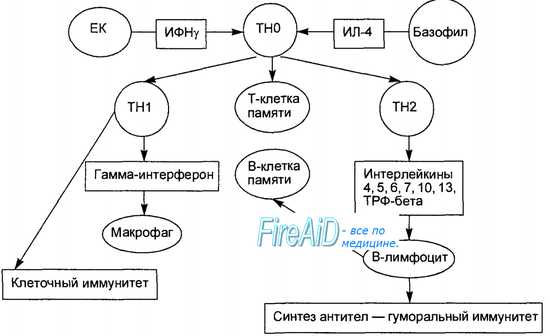 Передача сигнала активации от антигенраспознающего рецептора в ядро клетки приводит к активации конкретных генов, ответственных за митотическую активность клеток, контролирующих продукцию клеткой биологически активных молекул, участвующих в антимикробной защите (антитела, цитокины). В-лимфоцит получает одновременно два сигнала активации: от антигенраспознающего рецептора при его соединении с антигеном и от связывания его поверхностных костимулирующих молекул с соответствующими лигандами на Т-лимфоцитах. После этого В-лимфоцит пролиферирует и потомки его превращаются в зрелые антителопродуцирующие плазматические клетки. Т-лимфоцит в ответ на контакт с антигеном начинает пролиферировать, потомки его приобретают способность продуцировать определенные цитокины или превращаются в зрелые цитотоксические клетки. В зависимости от того, какие дополнительные сигналы активации (цитокины, костимулирующие молекулы) получает Т-лимфоцит в момент контакта с антигеном, его потомки дифференцируются в двух разных направлениях, превращаясь в Т-хелперы, продуцирующие гамма-интерферон (Th1), или в Т-хелперы, продуцирующие интерлейкины-4, -5, -6, -10, -13 (Th2). Количественное преобладание Thl над Th2 является условием развития клеточного (клеточно-опосредованного) иммунного ответа. В случае преобладания Th2 развивается гуморальный иммунный ответ, проявляющийся продукцией специфических антител (рис. 8.9). 5.Гуморальный и клеточный иммунитет Гуморальный иммунитет Гуморальный иммунитет связан с образованием антител. Антигенраспознающие рецепторы В-лимфоцитов представляют собой молекулы иммуноглобулинов. При связывании антигена с соответствующим рецептором и под влиянием цитокинов, вырабатываемых моноцитами, макрофагами и Т-лимфоцитами, происходит активация В-лимфоцитов, которые начинают делиться и дифференцироваться в лимфатических узлах в плазматические клетки(см. рис. ). Часть активированных В-лимфоцитов превращаются в клетки памяти, которые обеспечивают более быстрый и эффективный иммунный ответ при повторном контакте с антигеном. Выделяют 4 стадии первичного иммунного ответа. На первой стадии, которая занимает 3—4 сут, антитела к соответствующему антигену в сыворотке отсутствуют. На второй стадии появляютсяIgM, и спустя 10—14 сут после контакта с антигеном — IgG. На третьей стадии уровень антител остается постоянным. Четвертая стадия первичного иммунного ответа обычно растягивается на месяцы. Она характеризуется постепенным снижением уровня антител. Вторичный иммунный ответ развивается при повторном контакте с антигеном. Антитела, главным образом иммуноглобулины IgG, появляются быстрее и в более высоком титре, чем при первичном иммунном ответе. Клеточный иммунитет Клеточный иммунитет связан с образованием специализированных клеток, реагирующих с антигеном посредством его связывания и последующего разрушения (см. рис.). Иммунная реакция опосредована клетками - цитотоксическими Т-лимфоцитами и Т-хелперами. Цитотоксические Т-лимфоциты непосредственно контактируют с чужеродными клетками и разрушают их, а Т-хелперы вырабатывают биологически активные вещества — цитокины, активирующие макрофаги. По способности вырабатывать разные цитокины и участвовать в регуляции клеточного и гуморального иммунитета Т-хелперы подразделяются на Тh1 и Th2. Первые вырабатывают интерферонγ и интерлейкин-2, стимулируют пролиферацию цитотоксических Т-лимфоцитов и активируют макрофаги, вторые вырабатывают интерлейкины-4, -5, -6, стимулируют пролиферацию и дифференцировку В-лимфоцитов, а также синтез антител разных классов. Т-клетки связывают антигены, если они ассоциированы с расположенными на поверхности животных клеток определенными антигенными структурами, которые называются главным комплексом гистосовместимости. Главный комплекс гистосовместимости (ГКГС) - это группа генов и кодируемых ими антигенов клеточной поверхности, которые играют важнейшую роль в распознавании чужеродного и развитии иммунного ответа. Антигены ГКГС подразделяются на антигены классов I иII. Антигены ГКГС класса 1 необходимы для распознавания трансформированных клеток цитотоксическими Т-лимфоцитами. Важнейшая функция антигенов ГКГС класса II — обеспечение взаимодействия между Т-лимфоцитами и макрофагами в процессе иммунного ответа. Т-хелперы распознают чужеродный антиген лишь после его переработки макрофагами, соединения с антигенами ГКГС класса II и появления этого комплекса на поверхности макрофага. Способность Т-лимфоцитов распознавать чужеродные антигены только в комплексе с антигенами ГКГС называютограничением по ГКГС. Клеточный иммунитет проявляется в виде: 1. Аллергических реакций замедленного типа (например, туберкулиновые пробы), аллергический контактный дерматит. 2. Защиты против внутриклеточных паразитов. 3. Противовирусного и противогрибкового иммунитета. 4. Отторжения трансплантата. 5. Противоопухолевого иммунитета. NB! При иммунном ответе обычно действуют механизмы как гуморального, так и клеточного иммунитета, но в разной степени. Гуморальный иммунный ответ более быстрый, чем клеточный. Как в гуморальной, так и в клеточной системе вторичные иммунные ответы протекают быстрее и интенсивнее, чем первичные. 6. Что такое клональность лимфоцитов Клональность лимфоцитов – это свойство Т- и В- лимфоцитов в процессе дифференцировки перестраивания генов, кодирующих рецепторы для антигенов, в результате которого каждая клетка экспрессирует рецептор, уникальный по специфичности, а все потомки этой клетки (клоны) имеют рецепторы такой же специфичности. В процессе селекции погибает большинство опасных аутоспецифических клонов как Т-, так и В-клеток. Популяции Т- и В-лимфоцитов участвуют в иммунных реакциях клонального типа, при которых в ответ вовлекаются только клетки клонов, экспрессирующих рецепторы нужной специфичности (в отличие от естественных киллеров, не отличающихся друг от друга по специфичности). Клетки клонов отличаются друг от друга по специфичности рецепторов, следовательно, специфичностью распознавания и спектром функций. Молекулы патогенна, антигена и стрессорных клеток распознаются тремя типами рецепторов клеток иммунной системы. 7. Отличие клонов друг от друга Клон лимфоцитов – группа лимфоцитов, имеющих антигенраспознающие рецепторы одной специфичности, т.е. способные распознавать одну АГ детерминанту. 1 клон = 1 АГ-распознающий рецептор! В иммунной системе формируется множество клонов лимфоцитов способных распознавать огромное количество вариантов антигенных молекул, составляющих антигенный репертуар. Распознавание антигена рецепторами В лимфоцитов – приобретенный иммунитет. Реакция требует недель. Есть память. Антиген и антитела - белки. Антитела – это заякоренные рецепторы В-лимфоцитов. Они могут отщепляться и находиться виде растворимых рецепторов. Белковые молекулы с высокой афинностью распознают друг на друге некие специальные поверхности, образуются высокоафинные комплексы. В этом смысл этого принципа, которым руководят В-клетки. 8. Основные рецепторы Т, В, НК лимфоцитов, их роль в иммунологических процессах Т-клеточные рецепторы для антигенов представляют собой антителоподобные гетеродимеры, состоящие из полипептидных α- и β-цепей. Каждая из цепей имеет длину в 280 аминокислот, большая внеклеточная часть каждой цепи свернута в два Ig-подобных домена: один вариабельный (V) и один константный (С). Антителоподобный гетеродимер кодируется генами, которые собираются из нескольких генных сегментов в процессе развития Т-клеток в вилочковой железе. Несмотря на то, что Т- и В-клетки довольно легко идентифицировать по поверхностным маркерам (Т3 или CD3 на Т-клетках и поверхностные Ig на Вклетках), следует иметь также представление о наиболее важных дифференцировочных антигенах Т-лимфоцитов человека. Важнейшими из них являются: 1. CD2 (от англ. Claster of differentiation – кластер дифференцировки) – это антиген, обнаруживаемый на всех зрелых периферических Т-лимфоцитах (идентичен "рецептору эритроцитов барана", именно он обеспечивает образование розеток с эритроцитами барана – методика выявления Т-клеток). CD2 принимает участие в процессе неспецифической активации Т-клеток, что играет важную роль при созревании клеток в тимусе, т.к. пролиферация тимоцитов индуцируется до начала экспрессии специфического антигенного процесса. 2. CD3 – это мембраносвязанный белковый комплекс, состоящий из пяти гликопротеинов, связанный с антигенспецифическим рецептором (Ti). Этот комплекс "CD3+Ti" и представляет собой антигенспецифический Т-клеточный рецептор периферических Т-лимфоцитов человека. Связывание антигена, ассоциированного с детерминантами МНС, является специфическим сигналом для активации зрелой Т-клетки. При этом CD3 участвует в передаче сигнала внутрь клетки. Непосредственным результатом связывания антигена с рецептором является поступление в клетку ионов Са2+. 3. CD4 – антиген гликопротеиновой природы, который экспрессирует примерно на 2/3 периферических Т-лимфоцитов. На этапе созревания клеток в тимусе CD4 экспрессируется всеми клетками, а в ходе их дифференцировки сохраняется только на субпопуляции, переставшей экспрессировать CD8-антиген. В периферической крови примерно 5% клеток несут одновременно маркеры CD4 и CD8. Зрелые CD4+-Т-клетки включают Т-лимфоциты, функционально характеризуемые как хелперы и индукторы. При контакте Т-лимфоцитов (Ti/h – индукторов хелперов) с антигенпрезентирующей клеткой CD4 выступает в роли специфического места связывания детерминант белковых молекул МНС класса II. Особое значение имеет факт связывание молекулой CD4 оболочечных белков вируса иммунодефицита человека – возбудителя СПИД, что в результате эндоцитоза приводит к проникновению вируса внутрь клеток субпопуляции Ti/h. 4. CD8 – антиген, который экспрессируется примерно на 1/3 периферических Тклеток, созревающих из CD4+/CD8+-Т-лимфоцитов. Субпопуляция CD8+-Тклеток включает цитотоксические и супрессорные Т-лимфоциты. При контакте с клеткой-мишенью CD8 выступает в роли рецептора неполиморфных детерминант белков МНС класса I. 5. Антиген CD45R присутствует примерно на 50% Т-клеток (он экспрессируется также В-клетками и моноцитами). Клетки CD4+/CD45R идентифицированы как индукторы супрессоров, что дает возможность косвенно определять также функционально активные индукторы хелперов. 6. Антиген CD25 – гликопротеин, идентифицированный как низкоаффинный рецептор к интерлейкину-2 (IL-2). Совместно с белком 75К антиген CD25 образует высокоаффинный рецептор ИЛ-2. CD25 экспрессируется на активированных Т-лимфоцитах. Функции Т-лимфоцитов Регуляция иммунного ответа - обеспечивается специальными субпопуляциями клеток, активирующих и угнетающих иммунные реакции. Рецепторы В-лимфоцитов Рецептором B-лимфоцитов, узнающим антиген, является IgM. Мембраносвязанный IgM (mIgМ), как правило, представляет собой мономерный иммуноглобулин, т.е. отдельную единицу из четырех полипептидных цепей. Эта молекула имеет гидрофобную последовательность, расположенную на C-концевом участке тяжелой цепи и предназначенную для фиксации молекулы на клеточной мембране. Число молекул рецептора достигает 10 - 100тыс. на клетку. Главной функцией В-лимфоцитов также является специфическое распознавание антигена, которое ведет к их активации, пролиферации и дифференцировке в плазматические клетки -- продуценты специфических антител -- иммуноглобулинов, т.е. к гуморальному иммунному ответу. Чаще всего В-лимфоциты для развития гуморального иммунного ответа нуждаются в помощи Т-лимфоцитов в виде продукции активирующих цитокинов. NK клетки способствуют развитию адаптивного иммунного ответа через продукцию цитокинов и хемокинов 1 и 2 типа. Секреция этих факторов, активированными NK клетками, ведет не только к созреванию ДК, но и к дифференцировке Т- и В-лимфоцитов. Недавно в экспериментах Zingoni А. и коллег было выявлено, что активированные NK клетки способствуют TCR-зависимой пролиферации покоящихся аутологичных СD4'-Т-клеток под воздействием костимулирующих молекул, суперсемейства иммуноглобулинов и TNF. Эти данные раскрывают новую связь между врожденным и адаптивным иммунитетом. 9.Корецепторы рецепторы лимфоцитов, их роль иммунологических процессах. Корецепторы Т-лимфоцитов Это СD8 и CD4. Это дополнительные молекулы, которые находятся рядом с Т клеточным рецептором и распознают только МНС. Молекулы МНС бывают двух классов. На всех клетках организма есть МНСI класса. Корецептор к нему – это СD8. Молекулы МНСII находятся на поверхности антиген-презентирующих клеток (это дендритные клетки). Именно дендритная клетка, активированная в участке воспаления, будет нести много МНСII, на которой будет много элементов чужеродных пептидов. Такая клетка будет представлять этот комплекс Т-хелперам. Ко-рецептор на его поверхности – это молекула СD4. Исключение, при котором не требуется распознавание пептидов – это суперантигены. Это белки вирусного или бактериального происхождения, которые взаимодействуют с Т-клеточным рецептором и МНС, обеспечивая прочный контакт вне зависимости от того какой пептид там находится. Они активируют до 20% всех Т клеток, что приводит к чрезмерной продукции цитокинов и подавлению адаптивного иммунитета. Пример – стафилокковый энтеротоксин. Наряду с проявлением усиливающих эффектов, корецепторы способны передавать супрессорные сигналы при распознавании антигена и последующей активации клеток. Супрессорный эффект проявляется, например, при изолированном перекрестном «сшивании» молекул корецепторов моноклональными антителами. На фоне такого связывания стимуляция через TCR вызывает апоптоз Т-лимфоцитов. 10. Дифференцировка Т и В лимфоцитов, где происходит Полипотентная гемопоэтическая стволовая клетка (ГСК) - малодифференцированный клеточный элемент, способный к самоподдержанию, пролиферации и дифференцировке во все клетки крови. ГСК локализуются в красном костном мозге (концентрация до 1% среди мононуклеарных клеток), печени эмбриона (до 1%), ГСК циркулируют в пуповинной крови (до 1%), • периферической крови (около 0,1% от всех ядерных клеток)  Основные этапы дифференцировки В-лимфоцитов · СКК – стволовая кроветворная клетка костного мозга; · ЛСК – лимфоидная стволовая клетка (общий предшественник лимфопоэза); · пВ – предшественник В-лимфоцитов; · В0–лимфоциты; · В1 – лимфоциты (наивные В-лимфоциты); · В2 – лимфоциты; В эмбриогенезе человека первые В-лимфоциты появляются в печени и затем в костном мозге. После рождения В-клеточный иммунопоэз происходит в основном в костном мозге, где В2-лимфоциты проходят несколько последовательных стадий: лимфоидная клетка-предшественник —> ранняя про-В-клетка —> поздняя про-В-клетка большая пре-В-клетка -> малая пре-В-клетка -» незрелая В-клетка зрелая В-клетка. Основным процессом дифференцировки В-лимфоцитов является формирование антигенраспознающего В-клеточного рецептора! В процессе развития, дифференцировки и функционирования судьба В-клетки определяется количеством и качеством сигналов, получаемых через ВСК. Так же, как в тимусе, в отношении Т-лимфоцитов в костном мозге после стадии общего лимфоидного предшественника запускается процесс перегруппировки генов иммуноглобулинов. В процессе дифференцировки Т-лимфоцитов выделяют два основных этапа (как Вы помните, такие же два этапа выделяют в процессе дифференцировки В-лимфоцитов): 1. Антигеннезависимая дифференцировка – происходит постоянно в тимусе. 2. Антигензависимая дифференцировка – происходит в периферических органах иммунной системы только при контакте Т-лимфоцита с антигеном. 11. Популяции В-лимфоцитов В популяции В-лимфоцитов различают несколько субпопуляций: 1. В1-лимфоциты – предшественники плазмоцитов, синтезирующих антитела без взаимодействия с Т- хелперами. Есть тимуснезависимые антигены (бактериальные полисахариды, полимеризованный флагелин, леван и др.), которые способны без лимфоцитов реагировать, т.е. фиксироваться на рецепторах В-клеток. Эти антигены стимулируют синтез только Ig М. 2. В2 –лимфоциты, превращаются после антигенной стимуляции в плазмоциты с помощью Т-хелперов, ответственны за гуморальный ответ на тимусзависимые антигены, сопровождающийся синтезом иммуноглобулинов всех классов. 3. В3-лимфоциты (В-киллеры) оказывают цитотоксическое действие на клетки-мишени, покрытые антителами, без участия комплемента. Допускается, что В-киллеры – производные «нулевых» лимфоцитов – лимфоцитов без отличительных признаков Т- и В- клеток. Тот факт, что они встречаются среди лимфоцитов костного мозга в 50 % случаев, а среди лимфоцитов крови в 5% случаев, позволяет предположить, что это незрелые формы лимфоцитов, хотя и обладающие цитотоксической активностью. 4. В-супрессоры тормозят пролиферацию и трансформацию Т-клеток, стимулированных антигеном. Супрессорное действие В-клеток, как и Т-клеток, осуществляется при непосредственном контакте с иммунокомпетентными клетками и опосредованно через медиаторы. 5. В-лимфоциты памяти формируются в ходе иммунного ответа на антиген, составляют около 1 % всех В-лимфоцитов, отличаются долголетием и способностью быстро отвечать на повторное поступление антигена. В- лимфоциты памяти не имеют морфологических отличий от других В -лимфоцитов, но обладают активным геном (bcl-2). В-клетки памяти рециркулируют между кровью, лимфой и лимфоидными органами, но более всего накапливаются в периферических лимфоидных органах. Они сохраняют информацию об антигене, способны передавать её другим клеткам, обеспечивают при повторном попадании антигена синтез Ig по вторичному признаку. 12. В2 и В1 – лимфоциты Они отличаются друг от друга тем, что выделяют разные группы антител. В1-лимфоциты выделяют антитела, которые характерны для борьбы с недавно проникшим в тело агрессором (иммуноглобулины М). Других защитных веществ они синтезировать не могут. По этой причине клетки данного вида располагаются в прибарьерных полостях. Они располагаются там затем, чтобы встретить микробы, когда те только-только прошли через защитные препятствия. По выполняемым задачам В1-лимфоциты можно сравнить с пограничниками: они ловят и устраняют только объекты, которые только что нарушили запретные барьеры. В2-клетки выделяют защитные факторы, которые активны преимущественно против инфекций, уже успевших обосноваться в организме (иммуноглобулины G). Их действия можно сравнить с активностью полиции, которая борется с действующими преступниками. Большая часть В-лимфоцитов в организме представлена В1-типом. 13. Основное свойство В-лимфоцитов Основное свойство В-лимфоцитов — экспрессия иммуноглобулинового рецептора для распознавания антигенов — BCR. На поверхности зрелой В-клетки содержится около 150 000 комплексов BCR. 14. Виды презентации антигена В-лимфоцитам Антигенраспознающий В-клеточный рецептор B-лимфоцитов (BсR – англ. В-cell Receptor) построен из молекулы мембранного иммуноглобулина (mIg, состоящий из двух одинаковых тяжелых H- и двух одинаковых легких - L-цепей) и двух молекул CD79 (Iga, Igb). BсR имеет трансмембранные и внутрицитоплазматические сегменты, передающие внутриклеточные сигналы.  15. Мембранные рецепторы В-лимфоцитов (основные, корецепторы) - перечислить B-лимфоциты экспрессируют следующие молекулы: 1)Антигенраспознающий B-клеточный рецептор (BсR) представленный, гл. обр., мембранными иммуноглобулинами – мономерами mIg M, mIg D; 2)Корецепторный комплекс мембранных молекул [CD19/CR2 (CD21)/TAPA-1], связанных с системами внутриклеточного проведения сигналов; 3)BсR-ассоциированные молекулы [Iga (CD79a) и Igb (CD79b)], необходимые для сигнальной трансдукции; 4)Костимулирующие молекулы (CD28, CD40 и др.) для дополнительных стимулов и переключения синтеза разных изотипов антител; 5)Адгезивные молекулы (ICAM-3 и др.) для контакта клеток. Основные маркеры В-лимфоцитов -мембранные Ig при этом клетки одного клона (быстро формирующегося в результате серии последовательных делений потом-ства одной В-клетки ) экспрессируют молекулы Ig, специфически связывающие только один эпитоп Аг. Такие клетки синтезируют моноклональные AT, способные распознавать и связывать только один Аг. Аг-связывающий участок мембранного Ig В-лимфоцита играет роль клеточного Аг-распознающего рецептора. Помимо мембранных Ig, В-лимфоцит несёт другие маркёры; рецепторы Fc-фрагмента Ig, CD10 {на незрелых В-клетках), CD19, CD20, CD21, CD22, CD23 (вероятно, участвуют в клеточной активации), рецепторы к С3b и C3d, молекулы МНС классов I и II. 16. Что такое Igα и Igβ, какую роль играют. Igα и Igβ - маленькие иммуноглобулиновые домены, ковалентно связанные между собой, имеющие внутриклеточный домен, в котором присутствует белковый мотив - ITAM. Это белковая последовательность, содержащая тирозин, фосфорилирование которой является сигнальным событием. Короткая цитоплазматическая часть BCR (три аминокислотных остатка) не может служить проводником внутриклеточного сигнала, индуцируемого взаимодействием BCR-антиген, поскольку не способна формировать молекулярные структуры, необходимые для взаимодействия с внутриклеточными белками. 17. Пути активации В-лимфоцитов Активация В-лимфоцитов связана с рядом специфических и неспецифических стимулов, которые приводят к глубоким биохимическим и морфологическим изменениям в этих клетках. 1. Активация В-клеток обусловлена внедрением липофильного блока в двойной липидный слой их мембраны. Это внедрение может происходить и без участия антигена, неспецифически, при большой концентрации соответствующего метаболита. Таким образом, например, действует, по-видимому, неспецифический стимулирующий фактор Т-клеток-помощников. Специфичность иммунологического процесса обусловлена наличием медиатора, который секретируется Т-лимфоцитами под влиянием антигена. Этот медиатор (секретируемый рецептор Т-клеток) состоит из трех частей: рецептора Т-клеток, специфически реагирующего с детерминантами несущей части антигена (носителя), Н-2-белка и «липофильного хвоста». Медиатор присоединяется своим рецептором к антигену, фиксированному на поверхности антигенсвязывающей клетки, и липофильный хвост внедряется в фосфолипидный слой мембраны. 2. Присоединение антигена к В-лимфоциту приводит к его обратимому параличу или инактивации благодаря возникновению сигнала 1. Сигнал 1 развивается при бимолекулярной реакции между одновалентным антигеном и соответствующим рецептором В-клетки. Он чрезвычайно быстро (возможно, уже через секунду) оказывает на клетку парализующее действие. Индукция иммунной реакции возникает в том случае, если на В-клетку после сигнала 1 подействует еще и сигнал 2. Первоначально предполагалось, что сигнал 2 вызывают антитела, которые фиксированы на поверхности Т-клеток и специфичны для антигена, присоединившегося к В-лимфоцитам. В дальнейшем была допущена возможность передачи сигнала 2 на небольшое расстояние при помощи образуемого Т-лимфоцитами медиатора. В некоторых случаях сигнал 2 обусловлен действием антител не против антигенов, присоединившихся к поверхности В-лимфоцитов, а против собственных антигенов его поверхности. 3. Для активации В-лимфоцитов необходим второй сигнал. Этот сигнал исходит от присоединившегося к клетке С3-компонента комплемента. В пользу этого взгляда говорят наличие на поверхности ПАОК рецепторов для активизированного С3-компонента и митогенность очищенных препаратов С3-компонента. Соответствует этой точке зрения и наличие в лимфатических клетках липосомальных протеаз, активирующих С3-компонент, и то, что эти протеазы выделяются как при действии на лимфатические клетки антигенов и иммуногенов, так и при взаимодействии Т- и В-лимфоцитов (Dukor, Hartman, 1973; Hartman, 1975). В связи с этой точкой зрения нельзя не упомянуть о митогенном действии протеаз. Действие на клетки селезенки трипсина стимулирует включение 3Н-тимидина в эти клетки почти так же сильно, как действие самых мощных митогенов. Действие трипсина направлено на В-клетки: более 80% трансформированных им клеток содержало на поверхности иммуноглобулины; он чрезвычайно сильно стимулирует включение 3Н-тимидина в клетки селезенок бестимусных мышей (Kaplan, Bona, 1974; Vischer, 1974). 4. По модели одного неспецифического сигнала присоединение антигена к иммуноглобулиновым рецепторам В-лимфоцитов индуцирует ряд процессов (например, образование «колпачка»), но не активирует эти клетки. В-лимфоциты активируются неспецифическим сигналом от участка на их поверхности, который не является иммуноглобулином. Эти сигналы исходят от самого антигена (но не от их антигенной детерминанты) в случае тимуснезависимых антигенов и от медиаторов Т-клеток или макрофагов (в случае тимусзависимых антигенов). 18.Особенности активации В-лимфоцитов путем перекрестной сшивки. Для эффективной активации В-клетки через BCR необходима перекрёстная «сшивка» антигеном нескольких BCR. Для этого молекула антигена должна иметь повторяющиеся эпитопы на своей поверхности. Эта сшивка захватывает соседние молекулы. 19.Что такое АПК Антигенпрезентирующие клетки Это гетерогенная популяция клеток, способных захватывать внеклеточные АГ, перерабатывать их в кислых условиях до пептидов и в ассоциации с молекулами МНС 2 класса представлять т-лимфоцитам для запуска адаптивного иммунного ответа К ним относятся макрофаги, дендритные клетки и В- лимфоциты 20. За счет чего В-лимфоциты являются АПК B-лимфоциты продуцируют и секретируют в кровоток молекулы антител, являющиеся измененными формами антигенраспознающих рецепторов этих лимфоцитов. Каждая B-клетка генетически запрограммирована на синтез молекул иммуноглобулина, встроенных в цитоплазматическую мембрану. Иммуноглобулины функционируют как антигенраспознающие рецепторы, специфичные к определенному антигену. 21. MHC – перечислить классы. Обнаружено две группы антигенов MHC (антигенов MHC класса I и антигенов MHC класса II ), участвующих в регуляции иммунного ответа. Эти группы антигенов по-разному экспрессируются на клетках организма и, хотя они выполняют однотипную функцию, между ними имеется "распределение обязанностей". Антигены MHC класса I представляют собой антигены, синтезируемые самой клеткой (вирусные, опухолевые, собственные мутированные), в то время как антигены MHC класса II - это экзогенные (пришедшие извне) антигены. 22. MHC 1, MHC 2 – функции. Основные функции молекулы MHC I – связывание антигенов (пептидов) и представление Т-лимфоцитам (Т-киллерам, ЦТЛ), α1 и α2 – домены при взаимодействии образуют удлиненную полость (щель), в которую встраивается антиген (пептид) из 8-10 аминокислот. В таком виде молекула способна мигрировать на поверхность клетки, где она выполняет свои функции. Этот комплекс является стабильным, что позволяет Т-киллеру сканировать антиген. На α3 домене есть участок связывания с молекулой CD8 Т-киллера. Важнейшая функция антигенов MHC (HLA) класса II - обеспечение взаимодействия между Т-лимфоцитами и макрофагами в процессе иммунного ответа. Т-хелперы распознают чужеродный антиген лишь после его переработки макрофагами , соединения с антигенами HLA класса II и появления этого комплекса на поверхности макрофага. 23. В1 – лимфоциты, где находятся, какие функции выполняют Образуются из стволовых клеток в костном мозге, обеспечивают гуморальный иммунитет. Из красного костного мозга мигрируют в В-зоны периферических лимфоидных органов, где под влиянием антигена дифференцируются в плазмоциты, вырабатывающие антитела (иммуноглобулины) и в В-клетки памяти. Функции лимфоцитов В-типа состоят в том, что они вырабатывают антитела – особые вещества, которые оказывают агрессивное воздействие на инородные частицы. Эти вещества растворимы и выделяются лимфоцитами в плазму крови. После того, как В-клетки выработали антитела против какого-то объекта, часть из этих лимфоцитов преобразуется в клетки памяти. Они формируют самую настоящую «библиотеку опасностей», хранят информацию о том, с чем столкнулись, и при повторной встрече с агрессором мобилизуют иммунитет на уничтожение «врага». 24.АПК клетки (профессиональные и непрофессиональные). Профессиональные антигенпрезентирующие клетки специализируются на представлении антигенов T-клеткам. Они очень эффективно поглощают антиген (по пути фагоцитоза, как макрофаги, или рецептор-опосредованного эндоцитоза, как B-клетки), разрезают его на пептидные фрагменты и затем экспонируют их на поверхности в составе комплекса с MHC-II. Комплекс MHC-II-антиген на мембране антигенпрезентирующей клетки распознаётся T-клетками с помощью T-клеточных рецепторов. Далее под действием дополнительного костимулирующего сигнала от АПК T-клетка активируется. Экспрессия костимулирующих молекул и MHC II класса — отличительная черта профессиональных антигенпрезентирующих клеток. Все профессиональные АПК, кроме того, также экспрессируют MHC I класса. Непрофессиональными АПК могут выступать все клетки организма, имеющие ядро. Они могут экспонировать эндогенные пептиды на своей поверхности с помощью молекулы MHC I класса и микроглобулина β-2. В отличие от пептидов чужеродного происхождения, которые экспонируют профессиональные АПК, непрофессиональные ЭПК экспонируют пептиды клеточного происхождения. T-киллеры могут распознавать эндогенные антигены в комплексе с MHC I класса. Как правило, непрофессиональные АПК не экспрессируют MHC II класса, хотя презентация антигенов в комплексе с MHC II класса не ограничена только профессиональными АПК. Другие лейкоциты, такие как нейтрофилы и тучные клетки, относящиеся к гранулоцитам, а также эпителиальные и эндотелиальные клетки при некоторых условиях могут экспонировать антигены в комплексе с MHC II класса. Однако убедительных доказательств того, что они могут активировать наивные CD4+ T-клетки, нет. |