03 подцарство многоклеточные. Подцарство Многоклеточные (Metazoa)
 Скачать 1.47 Mb. Скачать 1.47 Mb.
|

|
Подцарство Многоклеточные (Metazoa) У представителей этого подцарства тело состоит из множества клеток, выполняющих различные функции. В связи со специализацией клетки многоклеточных обычно теряют способность к самостоятельному существованию. Целостность организма обеспечивается путем межклеточных взаимодействий. Индивидуальное развитие, как правило, начинается с зиготы, характеризуется дроблением зиготы на множество клеток-бластомеров, из которых в дальнейшем формируется организм с дифференцированными клетками и органами. Филогения многоклеточных Происхождение многоклеточных от одноклеточных в настоящее время считается доказанным. Главным доказательством этого является почти полная идентичность структурных компонентов клетки многоклеточных животных структурным компонентам клетки простейших. Гипотезы происхождения многоклеточных подразделяются на две группы: а) колониальные, б) полиэргидные гипотезы. Колониальные гипотезы Сторонники колониальных гипотез считают, что переходной формой между одноклеточными и многоклеточными животными являются колониальные простейшие. Ниже перечисляются и кратко характеризуются гипотезы этой группы.
За переходную форму между одноклеточными и многоклеточными животными И.И. Мечников также принимает «бластею» (однослойная шаровидная колония жгутиковых). От «бластеи» происходят первые многоклеточные – «фагоцителлы». «Фагоцителла» не имеет рта, тело ее состоит из двух слоев клеток, жгутиковые клетки наружного слоя выполняют двигательную функцию, внутреннего – функцию фагоцитоза. «Фагоцителла» образуется из «бластеи» путем иммиграции части клеток наружного слоя внутрь колонии. Прообразом, или живой моделью гипотетического предка многоклеточных – «фагоцителлы» – И.И. Мечников считал личинку губок – паренхимулу.
Полиэргидные гипотезы Сторонники полиэргидных гипотез считают, что переходной формой между одноклеточными и многоклеточными животными являются полиэргидные (многоядерные) простейшие. По мнению И. Хаджи (1963), предками многоклеточных были многоядерные инфузории, первыми многоклеточными – плоские черви типа планарий. Наиболее аргументированной является гипотеза «фагоцителлы» И.И. Мечникова, доработанная А.В. Ивановым. Подцарство Многоклеточные подразделяется на три надраздела: 1) Фагоцителлообразные, 2) Паразои, 3) Эуметазои. Тип Пластинчатые (Placozoa) Трихоплакс Пластинчатые – самые примитивные из современных многоклеточных животных. К этому типу относятся лишь два вида одного рода морских животных – трихоплаксов. Трихоплакс (Trichoplax adhaerens) (см. рис.) – очень сильно уплощенное морское животное, обитающее на водорослях. Напоминает амебу, поскольку при движении контуры тела меняются. Движется при помощи жгутиковых клеток, покрывающих тело. Внутри тела располагаются веретеновидные и амебоидные клетки. 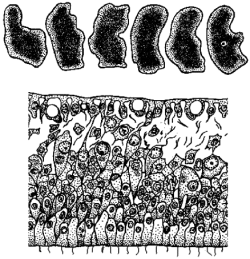 Вверху - изменения формы тела одной особи трихоплакса, зарисованные через каждые 10 мин. Внизу - поперечный разрез через тело трихоплакса. У трихоплакса обнаружено два способа питания: 1) внеклеточное, 2) фагоцитоз. При первом способе клетки «брюшной» стороны выделяют ферменты, которые переваривают бактерии, покрывающие водоросли. При втором способе пищевые частицы биением жгутиков загоняются на спинную сторону. Здесь эти частицы поглощаются веретеновидными клетками, которые выдвигаются наружу между жгутиковыми клетками. После поглощения пищевой частицы веретеновидная клетка становится амебоидной и погружается внутрь трихоплакса. Размножаются трихоплаксы бесполым и половым способами. Бесполое размножение осуществляется или путем деления тела надвое, или путем отпочковывания «бродяжек». Трихоплакс обладает высокой способностью к регенерации. Трихоплаксы, по-видимому, являются потомками первых многоклеточных животных на Земле. Их строение соответствует строению «фагоцителлы», гипотетического организма, предложенного И.И. Мечниковым для объяснения механизма происхождения многоклеточных животных. Тип Губки (Porifera, или Spongia) Строение и классы губок. Губки – древние примитивные многоклеточные животные. Обитают в морских, реже пресных водоемах. Ведут неподвижный прикрепленный образ жизни. Являются фильтраторами. Большая часть видов образует колонии. Не имеют тканей и органов. Почти все губки обладают внутренним скелетом. Скелет образуется в мезоглее, может быть минеральным (известковым или кремниевым), роговым (спонгиновым) или смешанным (кремниево-спонгиновым). Выделяют три типа строения губок: аскон (асконоидный), сикон (сиконоидный), лейкон (лейконоидный) (рис. 1).  рис. 1. Различные типы строение губок: 1 - аскон, 2 - сикон, 3 - лейкон. Наиболее просто организованные губки асконоидного типа имеют форму мешка, который основанием прикреплен к субстрату, а устьем (оскулумом) обращен кверху. Наружный слой стенки мешка образован покровными клетками (пинакоцитами), внутренний – воротничковыми жгутиковыми клетками (хоаноцитами). Хоаноциты выполняют функцию фильтрации воды и фагоцитоза. Между наружным и внутренним слоями располагается бесструктурная масса – мезоглея, в которой находятся многочисленные клетки, в том числе образующие спикулы (иглы внутреннего скелета). Все тело губки пронизано тонкими каналами, ведущими в центральную атриальную полость. Непрерывная работа жгутиков хоаноцитов создает ток воды: поры → поровые каналы → атриальная полость → оскулум. Питается губка теми пищевыми частицами, которые приносит вода. 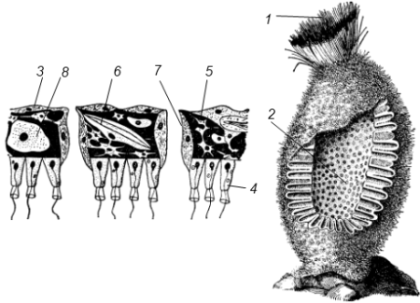 рис. 2. Строение сикона (Sycon sp.): 1 - скелетные иглы, окружающие устье, 2 - атриальная полость, 3 - пинакоцит, 4 - хоаноцит, 5 - звездчатая опорная клетка, 6 - спикула, 7 - пора, 8 - амебоцит. У губок сиконоидного типа происходит утолщение мезоглеи и образование внутренних впячиваний, имеющих вид карманов, выстланных жгутиковыми клетками (рис. 2). Ток воды в сиконоидной губке осуществляется по следующему пути: поры → поровые каналы → жгутиковые карманы → атриальная полость → оскулум. Наиболее сложный тип губок – лейкон. Для губок этого типа характерен мощный слой мезоглеи с множеством скелетных элементов. Внутренние впячивания погружаются вглубь мезоглеи и приобретают вид жгутиковых камер, соединяющихся выносящими каналами сатриальной полостью. Атриальная полость у лейконоидных губок, так же как у сиконоидных выстлана пинакоцитами. Лейконоидные губки обычно образуют колонии с множеством устьев на поверхности: в виде корок, пластинок, комьев, кустов. Ток воды в лейконоидной губке осуществляется по следующему пути: поры → поровые каналы → жгутиковые камеры → выносящие каналы → атриальная полость → оскулум. Губки обладают очень высокой способностью к регенерации. Размножаются бесполым и половым способами. Бесполое размножение осуществляется в форме наружного почкования, внутреннего почкования, фрагментации, образования геммул и пр. При половом размножении из оплодотворенной яйцеклетки развивается бластула, состоящая из одного слоя клеток, имеющих жгутики (рис. 3). Затем часть клеток мигрирует внутрь и превращается в амебоидные клетки. После того, как личинка оседает на дно, происходит перемещение жгутиковых клеток внутрь, они становятся хоаноцитами, а амебоидные клетки выходят на поверхность и превращаются в пинакоциты. 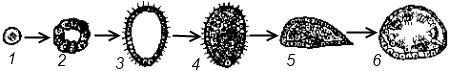 рис. 3. Развитие известковой губки (Clathrina sp.): 1 - зигота, 2 - равномерное дробление, 3 - целобластула, 4 - паранхимула в воде, 5 - осевшая паранхимула с инверсией пластов, 6 - молодая губка. Далее личинка превращается в молодую губку. То есть первичная эктодерма (мелкие жгутиковые клетки) становится на место энтодермы, а энтодерма – на место эктодермы: зародышевые пласты меняются местами. На этом основании зоологи называют губок животными, вывернутыми наизнанку (Enantiozoa). Личинка большинства губок – паренхимула, по строению почти полностью соответствует гипотетической «фагоцителле» И.И. Мечникова. В связи с этим в настоящее время наиболее обоснованной считается гипотеза происхождения губок от фагоцителлообразного предка. Тип Губки подразделяется на классы: 1) Известковые губки, 2) Стеклянные губки, 3) Обыкновенные губки. Класс Известковые губки (Calcispongiae, или Calcarea) Морские одиночные или колониальные губки с известковым скелетом. Скелетные иглы могут быть трех-, четырех- и одноосными. К этому классу относится сикон (рис. 2). Класс Стеклянные губки (Hyalospongia, или Hexactinellida)  рис. 4. Корзинка Венеры (Euplectella asper)  рис. 5. Туалетная губка (Spongia officianalis)  рис. 6. Кубок Нептуна (Poterion neptuni) Морские глубоководные губки с кремниевым скелетом, состоящим из шестиосных игл. У ряда видов иглы спаиваются, образуя амфидиски или сложные решетки. Скелеты некоторых видов очень красивы и используются в качестве коллекционных объектов и сувениров. Представители: корзинка Венеры (рис. 4), гиалонема. Класс Обыкновенные губки (Demospongiae) К этому классу относится подавляющее большинство современных видов губок. Скелет – кремниевый в сочетании со спонгиновыми нитями. У некоторых видов кремниевые иглы редуцируются, остаются лишь спонгиновые нити. Кремниевые иглы – четырех- или одноосные. Представители: туалетная губка (рис. 5), кубок Нептуна (рис. 6), бадяга, обитающая в пресных водоемах Тип Кишечнополостные (Coelenterata) Кишечнополостные обитают в морских, редко – в пресных водоемах. Образ жизни – планктонный или бентосный. Морфологические формы, приспособленные к планктонному образу жизни, называются медузами. Морфологические формы, приспособленные к бентосному образу жизни, называются полипами. Некоторые виды образуют колонии из полипоидных или медузоидных особей. Для кишечнополостных характерна радиальная симметрия. Клетки в их телах располагаются двумя слоями (двуслойные животные). Наружный слой клеток называется эктодермой, внутренний слой – энтодермой. Между эктодермой и энтодермой находится или тонкая базальная мембрана, или мезоглея – студенистый неклеточный слой. Все кишечнополостные имеют гастральный (напоминающий гаструлу) тип строения: представляют собой двухслойный «мешок» с полостью внутри. Эта полость тела открывается наружу одним ротовым отверстием и называется кишечной или гастральной. 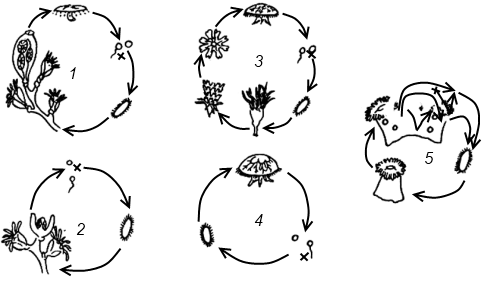 Схемы жизненных циклов кишечнополостных с чередованием поколений (1, 3) и без чередования поколений (2, 4, 5): 1, 2 - гидроидные полипы;) 3, 4 - сцифоидные полипы; 5 - коралловые полипы. В эктодерме произошла дифференциация клеток на эпителиально-мускульные, стрекательные, промежуточные (интерстициальные) и нервные. Наличие стрекательных клеток – специфический признак типа Кишечнополостные. Энтодерма подразделяется на эпителиально-мускульные и железистые клетки. Нервные клетки соединяются своими отростками, образуя нервную систему диффузного типа. Пищеварение происходит в гастральной полости и становится полостным, но сохраняется и внутриклеточное пищеварение, так как клетки энтодермы способны к фагоцитозу. Непереваренные остатки пищи удаляются из организма через ротовое отверстие. Железистые клетки секретируют в гастральную полость пищеварительные ферменты. Размножение – бесполое (почкование) и половое. Для многих кишечнополостных характерно чередование поколений (см. рис.). Тип подразделяется на три класса: 1) Гидроидные (Hydrozoa), 2) Сцифоидные медузы (Scyphozoa), 3) Коралловые полипы (Anthozoa). Кишечнополостные – древняя группа. В протерозойской эре выделяют огромный по продолжительности (90–100 млн. лет) вендский период, в фауне которого господствовали кишечнополостные. Сохранилось множество отпечатков медуз и бесскелетных полипов. Предполагается, что предками кишечнополостных были двухслойные плавающие многоклеточные, похожие на планулу. Вероятней всего, что первыми кишечнополостными были одиночные полипы, развивающиеся без смены поколений. Класс Гидроидные (Hydrozoa) Гидра. Обелия. Строение гидры. Гидроидные полипы Обитают в морских, редко – в пресных водоемах. Гидроидные – наиболее просто организованные кишечнополостные: гастральная полость без перегородок, нервная система без ганглиев, половые железы развиваются в эктодерме. Нередко образуют колонии. У многих в жизненном цикле имеется смена поколений: полового (гидроидные медузы) и бесполого (полипы) (см. Кишечнополостные). 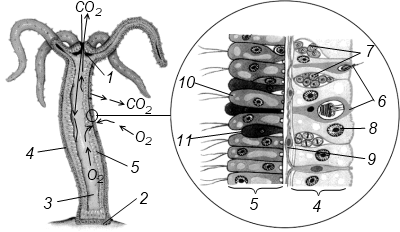 рис. 1. Гидра: 1 - рот, 2 - подошва, 3 - гастральная полость, 4 - эктодерма, 5 - энтодерма, 6 - стрекательные клетки, 7 - интерстициальные клетки, 8 - эпителеально-мускульная клетка эктодермы, 9 - нервная клетка, 10 - эпителеально-мускульная клетка энтодермы, 11 - железистая клетка. Гидра (Hydra sp.) (рис. 1) – одиночный пресноводный полип. Длина тела гидры около 1 см, нижняя его часть – подошва – служит для прикрепления к субстрату, на противоположной стороне находится ротовое отверстие, вокруг которого располагаются 6–12 щупалец. Как у всех кишечнополостных, клетки гидры располагаются двумя слоями. Наружный слой называется эктодермой, внутренний – энтодермой. Между этими слоями – базальная пластинка. В эктодерме выделяют следующие виды клеток: эпителиально-мускульные, стрекательные, нервные, промежуточные (интерстициальные). Из мелких недифференцированных интерстициальных клеток могут формироваться любые другие клетки эктодермы, в том числе в период размножения и половые клетки. В основании эпителиально-мускульных клеток находятся мускульные волокна, расположенные вдоль оси тела. При их сокращении тело гидры укорачивается. Нервные клетки имеют звездчатую форму и располагаются на базальной мембране. Соединяясь своими длинными отростками, они образуют примитивную нервную систему диффузного типа. Ответная реакция на раздражение имеет рефлекторный характер. В эктодерме присутствуют стрекательные клетки трех типов: пенетранты, вольвенты и глютинанты. Клетка-пенетрант – грушевидной формы, имеет чувствительный волосок – книдоциль, внутри клетки располагается стрекательная капсула, в которой находится спирально закрученная стрекательная нить. Полость капсулы заполнена токсичной жидкостью. На конце стрекательной нити находятся три шипика. Прикосновение к книдоцилю вызывает выброс стрекательной нити. При этом в тело жертвы вначале вонзаются шипики, затем по каналу нити впрыскивается яд стрекательной капсулы. Яд оказывает болевое и парализующее действие. Стрекательные клетки двух других типов выполняют дополнительную функцию удерживания добычи. Вольвенты выстреливают ловчие нити, опутывающие тело жертвы. Глютинанты выбрасывают клейкие нити. После выстреливания нитей стрекательные клетки отмирают. Новые клетки образуются из интерстициальных. Гидра питается мелкими животными: ракообразные, личинки насекомых, мальки рыб и др. Добыча, парализованная и обездвиженная с помощью стрекательных клеток, направляется в гастральную полость. Переваривание пищи – полостное и внутриклеточное, непереваренные остатки выводятся через ротовое отверстие. Гастральная полость выстлана клетками энтодермы: эпителиально-мускульными и железистыми. В основании эпителиально-мускульных клеток энтодермы находятся мускульные волокна, расположенное в поперечном направлении по отношению к оси тела, при их сокращении тело гидры сужается. Участок эпителиально-мускульной клетки, обращенный в гастральную полость, несет от 1 до 3 жгутиков и способен образовывать ложноножки для захвата пищевых частиц. Кроме эпителиально-мускульных имеются железистые клетки, секретирующие в кишечную полость пищеварительные ферменты. 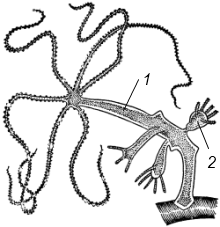 рис. 2. Почкование гидры: 1 - материнская особь, 2 - дочерняя особь (почка). Гидра размножается бесполым (почкование) и половым способами. Бесполое размножение происходит в весенне-летний сезон. Почки закладываются обычно на срединных участках тела (рис. 2). Через некоторое время молодые гидры отделяются от материнского организма и начинают вести самостоятельную жизнь. Половое размножение происходит осенью. В период полового размножения в эктодерме развиваются половые клетки. Сперматозоиды образуются на участках тела поблизости от ротового отверстия, яйцеклетки – ближе к подошве. Гидры могут быть как раздельнополыми, так и гермафродитными. После оплодотворения зигота покрывается плотными оболочками, образуется яйцо. Гидра погибает, а из яйца следующей весной развивается новая гидра. Развитие прямое без личинок. Гидра обладает высокой способностью к регенерации. Это животное способно восстанавливаться даже из небольшой отрезанной части тела. За процессы регенерации отвечают интерстициальные клетки. Жизнедеятельность и регенерация гидры были впервые изучены Р. Трамбле. Обелия (Obelia sp.) – колония морских гидроидных полипов (рис. 3). Колония имеет вид кустика и состоит из особей двух видов: гидрантов и бластостилей. Эктодерма членов колонии выделяет скелетную органическую оболочку – перидерму, которая выполняет функции опоры и защиты. 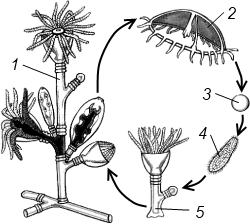 рис. 3. Жизненный цикл обелии: 1 - колония полипов, 2 - гидроидная медуза, 3 - яйцо, 4 - планула, 5 - молодой полип с почкой. Большая часть особей колонии – гидранты. Строение гидранта напоминает строение гидры. В отличие от гидры: 1) рот расположен на ротовом стебельке, 2) ротовой стебелек окружен множеством щупалец, 3) гастральная полость продолжается в общем «стебле» колонии. Пища, захваченная одним полипом, распределяется между членами одной колонии по разветвленным каналам общей пищеварительной полости. Бластостиль имеет вид стебелька, не имеет рта и шупалец. От бластостиля отпочковываются медузы. Медузы отрываются от бластостиля, плавают в толще воды и растут. Форму гидроидной медузы можно сравнить с формой зонтика. Между эктодермой и энтодермой находится студенистый слой – мезоглея. На вогнутой стороне тела, в центре, на ротовом стебельке находится рот. По краю зонтика свешиваются многочисленные щупальца, служащие для ловли добычи (мелкие рачки, личинки беспозвоночных и рыб). Число щупалец кратно четырем. Пища изо рта попадает в желудок, от желудка отходят четыре прямых радиальных канала, опоясывающие край зонтика медузы. Способ движения медузы «реактивный», этому способствует складка эктодермы по краю зонтика, называемая «парусом». Нервная система диффузного типа, но имеются скопления нервных клеток по краю зонтика. Четыре гонады образуются в эктодерме на вогнутой поверхности тела под радиальными каналами. В гонадах формируются половые клетки. Из оплодотворенной яйцеклетки развивается личинка паренхимула, соответствующая подобной личинке губок. Затем паренхимула преобразуется в двухслойную личинку планулу. Планула, поплавав при помощи ресничек, оседает на дно и превращается в нового полипа. Этот полип путем почкования образует новую колонию. Для жизненного цикла обелии характерно чередование бесполого и полового поколений. Бесполое поколение представлено полипами, половое – медузами. Класс Сцифоидные (Scyphozoa) Сцифоидные медузы: аурелия, цианея, корнерот Сцифоидные – кишечнополостные, специализированные к планктонному образу жизни. Большая часть жизненного цикла проходит в форме плавающих медуз, фаза полипа кратковременна или отсутствует. У сцифоидных медуз – такой же план строения, как у гидроидных медуз. В отличие от гидроидных, сцифоидные медузы имеют: 1) более крупные размеры, 2) сильно развитую мезоглею, 3) более развитую нервную систему с восемью обособленными ганглиями, 4) энтодермальные гонады, 5) желудок, разделенный на камеры. Способ передвижения – «реактивный», но поскольку сцифоидные не имеют «паруса», передвижение достигается за счет сокращения стенок зонтика. По краю зонтика располагаются комплексные органы чувств – ропалии. Каждый ропалий содержит «обонятельную ямку», орган равновесия и стимуляции движения зонтика – статоцист, светочувствительный глазок. Сцифоидные медузы – хищники, однако глубоководные виды питаются погибшими организмами. 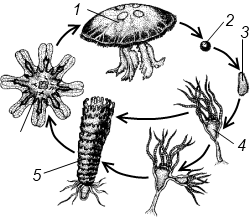 рис. 1. Жизненный цикл аурелии: 1 - взрослая особь, 2 - яйцо, 3 - планула, 4 - сцифистома, 5 - стробила, 6 - эфира. Аурелия (Aurelia aurita) (рис. 1) – одна из самых распространенных медуз. По краю зонтика располагаются мелкие щупальца. На вогнутой стороне в центре зонтика на коротком стебельке находится рот. Края рта вытянуты в четыре ротовые лопасти. Стрекательные клетки располагаются на щупальцах и ротовых лопастях. Желудок имеет четыре кармана, в которых имеются гастральные нити, увеличивающие пищеварительную поверхность. От карманов отходят восемь неветвящихся и восемь ветвящихся радиальных каналов. Радиальные каналы впадают в кольцевой канал. По неветвящимся каналам пища передвигается из желудка в кольцевой канал, по ветвящимся – в обратном направлении. По краю зонтика располагаются восемь ганглиев (скоплений нервных клеток), над ними – восемь ропалий. Ропалия представляет собой укороченное щупальце, внутри которого находится один статоцист, а по бокам – два глазка. На соседних укороченных щупальцах располагаются обонятельные ямки. Глазки обладают светочувствительной функцией.  рис. 2. Цианея (Cyanea arctica) Медузы – раздельнополые животные. Гонады образуются в энтодерме карманов желудка, имеют подковообразную форму. Созревшие половые клетки выделяются через рот медузы. Оплодотворение наружное. Яйца развиваются в складках ротовых лопастей. Внутри яйца формируется личинка планула. Планула покидает тело материнской особи. Проплавав некоторое время, планула опускается на дно и превращается в одиночного полипа – сцифистому. Сцифистома размножается почкованием, подобным почкованию гидры. Через некоторое время сцифистома преобразуется в стробилу, при этом щупальца сцифистомы укорачиваются, а на теле появляются поперечные перетяжки. Процесс поперечного деления называется стробиляцией. Путем стробиляции от стробилы отделяются молодые медузки – эфиры. Эфиры постепенно превращаются во взрослых медуз. 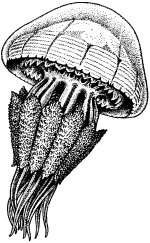 рис. 3. Корнерот (Rhizostoma pulmo) Цианея (Cyanea arctica) обитает в арктических морях. Является самой крупной медузой: диаметр зонтика может достигать 2 м, длина щупалец – 30 м (рис. 2). Цианея ярко окрашена, яд стрекательных капсул опасен для человека. Корнерот (Rhizostoma pulmo) не имеет щупалец по краям зонтика. Ротовые лопасти раздваиваются, их боковые стороны образуют многочисленные складки, которые срастаются между собой. Концы ротовых лопастей заканчиваются восемью корневидными выростами, от которых медуза получила свое название (рис. 3). Рот у взрослых корнеротов зарастает, пища поступает через многочисленные мелкие отверстия складок ротовых лопастей. Питается мелкими планктонными организмами. Встречается в Черном море. Ропилема съедобная (Rhopilema esculenta) наряду с аурелией употребляется в пищу в Китае и Японии. Ропилема напоминает черноморского корнерота, отличается от него желтоватой или красноватой окраской ротовых лопастей и наличием большого числа пальцевидных выростов. В пищу используется мезоглея зонтика. Класс Коралловые полипы (Anthozoa) Актинии. Коралловые рифы Коралловые полипы – морские колониальные, иногда одиночные полипы. По размерам коралловые полипы крупнее гидроидных. Тело имеет цилиндрическую форму, на верхней стороне тела располагается ротовое отверстие, окруженное щупальцами. Рот ведет в эктодермальную глотку. Кишечная полость разделена перегородками на камеры. Особенность коралловых полипов – наличие наружного (образованного эктодермой) или внутреннего (образованного мезоглеей) скелета. Отсюда – важнейшая роль коралловых полипов в образовании рифов. Встречаются коралловые полипы, лишенные скелета. Большая часть видов – раздельнополые животные. Гонады образуются в эндодерме. Размножение бесполое – почкованием, и половое – с метаморфозом, через стадию свободно плавающей личинки – планулы. Чередования поколений нет, поскольку медузоидная стадия отсутствует (см. Кишечнополостные).  рис. 1. Мягкий коралл дендронефтия Класс Коралловые полипы подразделяется на два подкласса: 1) Восьмилучевые кораллы (Octocorallia), 2) Шестилучевые кораллы (Hexacorallia). Подкласс Восьмилучевые кораллы (Octocorallia) У восьмилучевых кораллов восемь щупалец, восемь перегородок в гастральной полости, внутренний скелет. Этот подкласс подразделяется на отряды: 1) Альционарии (Alcyonaria), 2) Роговые кораллы (Gorgonacea) и др.  рис. 2. Благородный коралл Большая часть альционарий – мягкие кораллы, не имеющие выраженного скелета (рис. 1). Развитым известковым скелетом обладают только некоторые тубипоры. В мезоглее этих кораллов образуются трубочки, которые спаиваются друг с другом поперечными пластинками. Скелет по форме отдаленно напоминает орга´н, поэтому у тубипор есть еще одно название – органчики. Органчики участвуют в процессе образования рифов. Роговые кораллы, или горгонарии обладают внутренним роговым скелетом. К этому отряду относится красный, или благородный коралл (Corallium rubrum), являющийся объектом промысла (рис. 2). Из скелетов красного коралла изготавливают ювелирные изделия. Подкласс Шестилучевые кораллы (Hexacorallia)  рис. 4. Актиния телия (вид сверху)  рис. 3. Актиния метридиум У шестилучевых кораллов множество щупалец, число которых кратно шести. Гастральная полость разделена сложной системой перегородок, число которых также кратно шести. У большей части представителей имеется наружный известковый скелет, имеются группы, лишенные скелета. К подклассу Шестилучевые кораллы относятся отряды: 1) Актинии, 2) Мадрепоровые кораллы и др. Актинии – крупные одиночные полипы, лишенные скелета. Имеют самую разнообразную окраску, часто яркую, за что их называют морскими анемонами (рис. 3, 4). Могут медленно перемещаться на мускулистой подошве. Некоторые виды актиний вступают в симбиоз с раками-отшельниками. Рак-отшельник служит для актинии средством передвижения, а актиния своими щупальцами со стрекательными клетками защищает рака от врагов.  рис. 5. Мозговик диплория (часть шаровидной колонии) Мадрепоровые кораллы – как одиночные, так и колониальные полипы, для которых характерно наличие мощного известкового скелета. На больших глубинах (до 6000 м) обитают обычно мелкие одиночные формы, у берегов встречаются крупные полипы, а также ветвистые колонии (высотой до 1 м), образующие заросли – коралловые банки. Представители этого отряда – основные рифообразователи. К ним относятся мозговики (рис. 5), грибовидные кораллы и др. Коралловые рифы – образуются вследствие жизнедеятельности коралловых полипов, имеющих известковый скелет. В состав рифа входят главным образом мадрепоровые кораллы, частично – некоторые шестилучевые кораллы и другие животные, обладающие скелетом (моллюски, губки, мшанки). Рифообразующие кораллы обитают только в тропических областях Мирового океана, так как нуждаются в высокой и постоянной температуре воды, чувствительны косвещенности, солености воды и ее насыщенности кислородом. Зависимость распределения от освещенности обусловлена симбиозом коралловых полипов с одноклеточными водорослями (зооксантеллами). Рифы бывают трех типов: береговые, барьерные и атоллы. Атолл – коралловый остров кольцевидной формы. Согласно гипотезе Ч. Дарвина, исходным типом является береговой риф. Барьерные рифы и атоллы образуются в результате постепенного опускания суши (рис. 6). 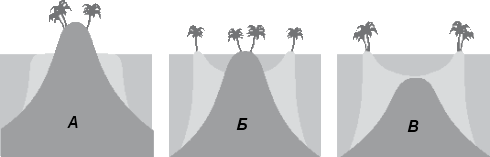 рис. 6. Происхождение коралловых рифов: А - береговой риф, Б - образование берьерного рифа из берегового, В - образование атолла из барьерного рифа. Тип Плоские черви (Plathelminthes) Плоские черви – двусторонне-симметричные трехслойные животные. Их тело – уплощенное в спинно-брюшном направлении и у большинства представителей имеет листообразную или лентовидную форму. Часть видов этого типа обитает в морских и пресных водоемах, большинство являются паразитами животных, в частности, человека. Покровы представлены кожно-мускульным мешком. У свободноживущих ресничных червей в состав кожно-мускульного мешка входят ресничный эпителий и три слоя мышц (кольцевые и продольные). У паразитических червей – тегумент и два слоя мышц. Кроме кольцевых и продольных мышц, у плоских червей имеются спинно-брюшные мышцы. Полость тела отсутствует, пространство внутри кожно-мускульного мешка между внутренними органами заполнено рыхлой массой соединительнотканных клеток – паренхимой. В промежутках между клетками циркулирует тканевая жидкость Нервная система состоит из парного головного ганглия и отходящих от него нескольких нервных стволов. Эти стволы соединяются друг с другом поперечными нервными тяжами (комиссурами). Органы чувств наиболее хорошо развиты у свободноживущих ресничных червей, которые имеют органы равновесия – статоцисты. Глаза, в отличие от глаз медуз, инвертированного типа. У всех плоских червей имеются рецепторы для восприятия механических и химических раздражений. Пищеварительная система присутствует у турбеллярии и сосальщиков, она состоит из двух отделов: переднего (эктодермального) и среднего (энтодермального). Кишечник слепо замкнут, задней кишки и анального отверстия нет. У цестод пищеварительная система отсутствует. 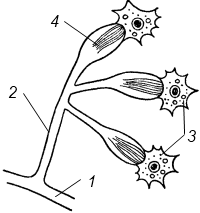 Схема строения протонефридиев: 1 - выводящий выделительный канал, 2 - разветвления канальцев, 3 - циртоциты ("звездчатые клетки"), 4 - реснички ("мерцательное пламя"). Выделительная система представлена отдельными клетками паренхимы (атроцитами) и протонефридиями. Протонефридии имеют эктодермальное происхождение и представляют собой систему ветвящихся каналов, выводящих из организма продукты жизнедеятельности в растворенном виде (см. рис.). Со стороны полости тела канальца замкнуты циртоцитами. Циртоцит – крупная клетка звездчатой формы, имеющая пучок ресничек («мерцательное пламя»). Биение ресничек обеспечивает отток межклеточной жидкости из паренхимы в протонефридиальный каналец. Канальца впадают в один или два выводящих канала, которые открываются наружу выделительными порами. В атроцитах происходит накопление продуктов жизнедеятельности. Специальные органы дыхания отсутствуют. Газообмен у свободноживущих и эктопаразитов (моногенеи) происходит через покровы тела. Эндопаразиты, живущие в среде, бедной кислородом, получают энергию за счет гликолиза. Кровеносная система отсутствует. Плоские черви гермафродиты. В состав половой системы, помимо семенников и яичников, входят придаточные образования, обеспечивающие процесс оплодотворения, снабжение яйцеклеток необходимыми питательными веществами и создание вокруг яйца защитных оболочек. Развитие плоских червей проходит в большинстве случаев с метаморфозом, через ряд личиночных стадий. Тип Плоские черви подразделяется на классы: 1) Ресничные черви (Turbellaria), 2) Сосальщики (Trematoda), 3) Ленточные черви (Cestoda) и др. Среди плоских червей только турбеллярии ведут свободный образ жизни, представители остальных классов являются специализированными паразитами. Поэтому вопрос о происхождении плоских червей сводится к выяснению происхождения турбеллярий. Считается, что предками турбеллярий являются фагоцителлообразные животные (гипотеза А.В. Иванова). В качестве первичных форм принимаются бескишечные турбеллярии (Acoela). От древних бескишечных турбеллярий произошли другие группы ресничных червей, в том числе прямокишечные турбеллярии (Rhabdocoela). Предполагается, что остальные классы плоских червей берут свое начало именно от древних прямокишечных турбеллярий. Переход к паразитизму осуществлялся, по-видимому, через симбиоз. Класс Ресничные черви (Turbellaria) Белая планария. Строение и органы Большая часть турбеллярий ведет свободный образ жизни, обитая в пресных и морских водоемах, влажных тропических лесах. В состав кожно-мускульного мешка входят однослойный ресничный эпителий и три слоя мышц. Рот обычно располагается на брюшной стороне тела. Строение пищеварительной системы у представителей разных отрядов различное: у одних видов кишечник может отсутствовать, у других – может быть достаточно сложным, разветвляющимся. Развитие у большинства видов – прямое, у морских многоветвистых турбеллярий – с метаморфозом. В последнем случае из яйца выходит «мюллеровская» личинка (рис. 1). Эта личинка имеет округлую форму, восемь радиально расположенных лопастей, покрыта ресничками. Лопасти помогают парить в толще воды. На брюшной стороне тела находится рот, ведущий в мешковидный кишечник. 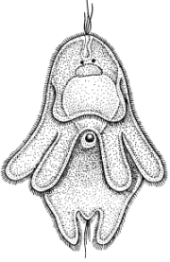 рис. 1. Мюллеровская личинка турбеллярий Класс Ресничные черви подразделяется на отряды: 1) Бескишечные (Acoela), 2) Многоветвистые (Polycladida), 3) Трехветвистые (Tricladida), 4) Прямокишечные (Rhabdocoela). Отряд Трехветвистые (Tricladida) 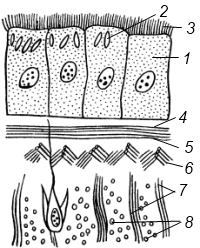 рис. 2. Схема строения кожно-мускульного мешка турбеллярий: 1 - клетки эпителия, 2 - рабдиты, 3 - реснички, 4 - базальная мембрана, 5 - колцевые мышцы, 6 - диагональные мышцы, 7 - спинно-брюшные мышцы, 8 - продольные мышцы. Представители этого отряда имеют «трехветвистый» кишечник. Половая система – сложная с хорошо развитыми желточниками. Развитие – прямое. Молочная, или белая планария (Dendrocoelum lacteum) достаточно часто встречается в пресных водоемах. Длина тела – до 2 см. Как у всех турбеллярий, в состав ее кожно-мускульного мешка входят однослойный ресничный эпителий и три слоя мышц (рис. 2). В эпителии находятся многочисленные железистые клетки, в частности, рабдитные клетки. Рабдитные клетки содержат палочковидные структуры – рабдиты. При раздражении они выбрасываются и образуют слизь, которая играет защитную функцию. Эпителий находится на базальной мембране, под которой располагается гладкая мускулатура: кольцевые, диагональные и продольные мышцы. Кроме этих, имеются пучки мышц, «протянутых» между спинной и брюшной сторонами тела: дорсовентральные мышцы. Молочные планарии передвигаются за счет биения ресничек и благодаря сокращениям мышц кожно-мускульного мешка. 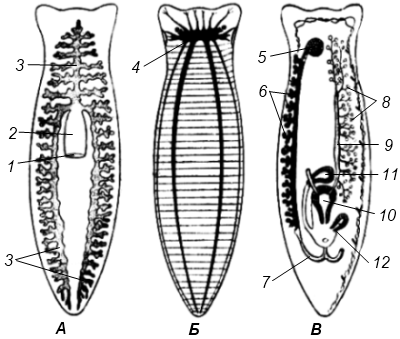 рис. 3. Схема строения молочной планарии. А - пищеварительная система, Б - нервная система, В - половая и выделительная системы: 1 - ротовое отверстие, 2 - глотка, 3 - ветви кишечника, 4 - нервные ганглии, 5 - яичник, 6 - желточники, 7 - яйцевод, 8 - семенные мешочки семенника, 9 - семяпровод, 10 - пенис, 11 - семяприемник, 12 - половая клоака. Ротовое отверстие у молочной планарии находится на вентральной стороне, ближе к задней части тела. Ротовое отверстие ведет в глотку, расположенную в глоточном кармане. От глотки отходят три ветви кишечника, одна из ветвей направлена к переднему концу тела, две другие – к заднему. Каждая ветвь имеет множество слепо замкнутых ответвлений второго порядка, за счет которых увеличивается площадь переваривания пищи (рис. 3А). Планария – хищник, питается мелкими беспозвоночными. Для захвата добычи используется мускулистая глотка, которая может выдвигаться из глоточного кармана. Пищеварение – внутриклеточное и кишечнополостное. В связи с отсутствием анального отверстия непереваренные остатки пищи выбрасываются через ротовое отверстие. Выделительная система, как у всех плоских, представлена протонефридиями (рис. 3В). В связи с обитанием в пресных водоемах, протонефридии молочной планарии развиты в гораздо большей степени, чем протонефридии морских видов. В состав нервной системы входят два нервных узла (ганглия) и отходящие от них нервные стволы (рис. 3Б). Ганглии расположены в передней части тела. Нервные тяжи соединяются друг с другом нервными комиссурами. Планария имеет два глазка инвертированного типа. Каждый глаз состоит из пигментного бокала, в его вогнутую часть погружены длинные зрительные клетки, на концах которых находятся светочувствительные структуры (рис. 4). Пигментный бокал расположен вогнутой частью к поверхности тела, так что свет сначала проходит через тела зрительных клеток, прежде чем попадет на световоспринимающие их участки. 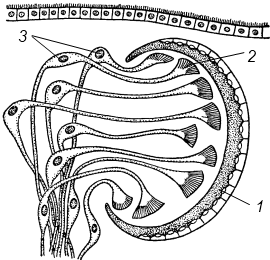 рис. 4. Схема строения глаза турбеллярии: 1 - пигментный бокал, 2 - светочувствителная |