Пособие по биологии для поступающих в вузы Авторы Пименов А. В., Гончаров О. В
 Скачать 34.81 Mb. Скачать 34.81 Mb.
|

38.2. Половое размножениеВ основе половогоразмножения лежит половой процесс, который связан с образованием большого количества специализированных клеток — гамет (половых клеток) и их последующего слияния. Сливаясь, гаметы образуют зиготы. Это приводит к уменьшению числа исходных клеток. Из зигот развиваются новые организмы, объединяющие в себе наследственную информацию родительских форм. Половое размножение характерно для большинства живых организмов. Для полого размножения характерны следующие особенности:
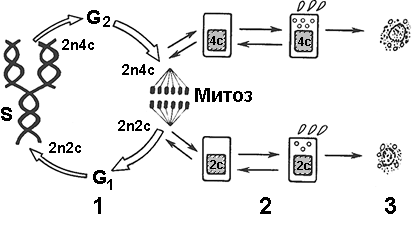
Как правило, яйцеклетки и сперматозоиды вырабатываются разными организмами. Такие организмы называются раздельнополыми. Если же один и тот же организм способен продуцировать и женские, и мужские гаметы, то его называют гермафродитом (ленточные черви, сосальщики). Но и в этом случае зигота образуется, чаще всего, в результате слияния гамет разных организмов (перекрестное оплодотворение). 38.3. Деление клетокВ основе передачи наследственной информации, размножения, развития, регенерации лежит деление клеток. Клетка как таковая существует только в промежутке между делениями. Жизненный (клеточный цикл)Период существования клетки от момента ее образования путем деления материнской клетки (включая само деление) до собственного деления или смерти называют жизненным (клеточным) циклом (рис. 305).В жизненном цикле клетки различают несколько фаз:
лением жизненных функций клетки и заканчивающийся ее делением или гибелью. Продолжительность жизненного цикла и количество составляющих его фаз у клеток различны. Так, клетки нервной ткани после завершения эмбрионального периода перестают делиться и функционируют на протяжении всей жизни организма, а затем погибают. Клетки же зародыша на стадии дробления, завершив одно деление, сразу же приступают к следующему, минуя все остальные фазы. Существует два способа деления клеток:
МитозМитоз20 — непрямое деление соматических клеток, представляющее собой непрерывный процесс, в результате которого сначала происходит удвоение, а затем равномерное распределение наследственного материала между дочерними клетками.Биологическое значение митоза:
Митотическое деление клетки находится под генетическим контролем. Митоз представляет собой центральное событие митотического цикла клетки. Митотический циклМитотический цикл — комплекс взаимосвязанных и детерминированных хронологически событий, происходящих в процессе подготовки клетки к делению и на протяжении самого деления.Длительность митотического цикла у разных организмов сильно варьирует. Самые короткие митотические циклы характерны для дробящихся яиц некоторых животных (например, у золотой рыбки первые деления дробления совершаются через 20 минут). Наиболее распространены митотические циклы длительностью 18-20 ч. Встречаются циклы продолжительностью несколько суток. Даже в пределах одного организма наблюдаются различия в продолжительности митотического цикла: клетки эпителия двенадцатиперстной кишки мыши делятся каждые 11 часов, тощей кишки — 19 часов, в роговице глаза — через 3 суток. Факторы, побуждающие клетку к митозу, точно не известны. Полагают, что основную роль играет соотношение объемов ядра и цитоплазмы (ядерно-цитоплазматическое соотношение). По некоторым данным, отмирающие клетки продуцируют вещества, способные стимулировать деление клетки. По двум главным событиям митотического цикла в нем выделяют:
Новые клетки появляются в ходе двух последовательных процессов:
Непосредственно на деление клетки уходит обычно 1-3 часа, то есть основную часть жизни клетка находится в интерфазе. ИнтерфазаИнтерфазой называют промежуток между двумя клеточными делениями. Продолжительность интерфазы, как правило, составляет до 90% всего клеточного цикла. Состоит из трех периодов:
Пресинтетический периодНачальный отрезок интерфазы — пресинтетический период (2n2c), период роста, начинающийся непосредственно после митоза. Самый длинный период интерфазы, продолжительность которого в клетках составляет от 10 часов до нескольких суток. Непосредственно после деления восстанавливаются черты организации интерфазной клетки:
Таким образом, в пресинтетический период осуществляются процессы подготовки следующего периода интерфазы — синтетического. Синтетический периодПродолжительность синтетического периода различна: от нескольких минут у бактерий до 6-12 часов в клетках млекопитающих.Во время синтетического периода происходит самое главное событие интерфазы — удвоение молекул ДНК. Каждая хромосома становится двухроматидной, а число хромосом не изменяется (2n4c). Параллельно с репликацией ДНК в цитоплазме интенсивно синтезируются гистоновые белки, которые затем мигрируют в ядро, где соединяются с ДНК. Постсинтетический периодНесмотря на то, что период называется постсинтетическим, это не означает отсутствие процессов синтеза на этом этапе интерфазы. Постсинтетическим его называют только потому, что он начинается после завершения синтеза (репликации) ДНК.Если пресинтетический период осуществлял рост и подготовку к синтезу ДНК, то постсинтетический обеспечивает подготовку клетки к делению и также характеризуется интенсивными процессами синтеза. В этот период:
Механизм митозаДеление ядра и цитоплазмы — это два самостоятельных процесса, проходящие непрерывно и последовательно. Однако для удобства изучения происходящих во время деления событий митоз искусственно разделяют на четыре стадии (рис. 306):
Длительность стадий митоза различна и зависит от типа ткани, физиологического состояния организма, внешних факторов. Наиболее продолжительны первая и последняя. Профаза(2n4c)Первая фаза деления ядра. В начале профазы (ранняя профаза) ядро заметно увеличивается. В результате спирализации хромосомы уплотняются, укорачиваются. В поздней профазе хорошо видно, что каждая хромосома состоит из двух хроматид, соединенных центромерой. Хромосомы начинают передвигаться к клеточному экватору.В поздней профазе из материала цитоплазмы формируется веретено деления. Оно образуется либо с участием центриолей (в клетках животных и некоторых низших растений), либо без них (в клетках высших растений и некоторых простейших). От центриолей, разошедшихся к разным полюсам клетки, начинают образовываться нити веретена деления двух типов:
К концу профазы ядерная оболочка исчезает, и хромосомы свободно располагаются в цитоплазме. Ядрышко обычно исчезает чуть раньше. Метафаза(2n4c)Началом метафазы считают тот момент, когда ядерная оболочка полностью исчезла. В начале метафазы хромосомы выстраиваются в плоскости экватора, образуя так называемую метафазную пластинку. Причем центромеры хромосом лежат строго в плоскости экватора. Нити веретена прикрепляются к центромерам хромосом, некоторые нити проходят от полюса к полюсу клетки, не прикрепляясь к хромосомам.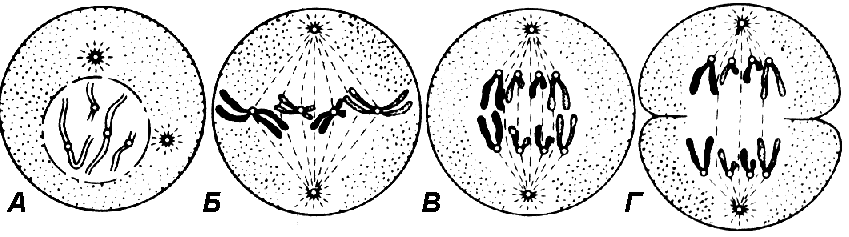 Рис. 306. Основные стадии митоза: А — профаза; Б — метафаза; В — анафаза; Г — телофаза. Анафаза(4n4c)Начинается с деления центромер всех хромосом, в результате чего хроматиды превращаются в две совершенно обособленные, самостоятельные дочерние хромосомы.Затем дочерние хромосомы начинают расходиться к полюсам клетки. Во время движения к полюсам они обычно принимают V-образную форму. Расхождение хромосом к полюсам происходит за счет укорачивания нитей веретена. В это же время происходит удлинение опорных нитей веретена, в результате чего полюса еще дальше отодвигаются друг от друга. Телофаза (2n2c)В телофазе хромосомы концентрируются на полюсах клетки и деспирализуются. Веретено деления разрушается. Вокруг хромосом формируется оболочка ядер дочерних клеток. На этом завершается деление ядра (кариокинез), затем происходит деление цитоплазмы клетки (или цитокинез).При делении животных клеток, на их поверхности в плоскости экватора появляется борозда, которая, постепенно углубляясь, разделяет материнскую клетку на две дочерние. У растений деление происходит путем образования так называемой клеточной пластинки, разделяющей цитоплазму. Она возникает в экваториальной области веретена, а затем растет во все стороны, достигая клеточной стенки (т.е. растет изнутри кнаружи). Клеточная пластинка формируется из материала, поставляемого эндоплазматической сетью. Затем каждая из дочерних клеток образует на своей стороне клеточную мембрану, и, наконец, на обеих сторонах пластинки образуются целлюлозные клеточные стенки. МейозМейоз — основной этап гаметогенеза, т.е. образования половых клеток. Образование гамет включает и митоз, и мейоз. Митоз обеспечивает накопление в половых железах большого количества незрелых клеток, которые впоследствии дают начало зрелым половым клеткам. Именно в результате мейоза происходит их созревание.Во время мейоза происходит не одно (как при митозе), а два следующих друг за другом клеточных деления. Первому мейотическому делению предшествует интерфаза I — фаза подготовки клетки к делению, в это время происходят те же процессы, что и в интерфазе митоза. Первое мейотическое деление называют редукционным, так как именно во время этого деления происходит уменьшение числа хромосом, то есть диплоидный набор хромосом становится гаплоидным, однако хромосомы остаются двухроматидными. Сразу же после первого деления мейоза совершается второе — обычный митоз. Это деление называют эквационным, так как во время этого деления хромосомы становятся однохроматидными. Биологическое значение мейоза:
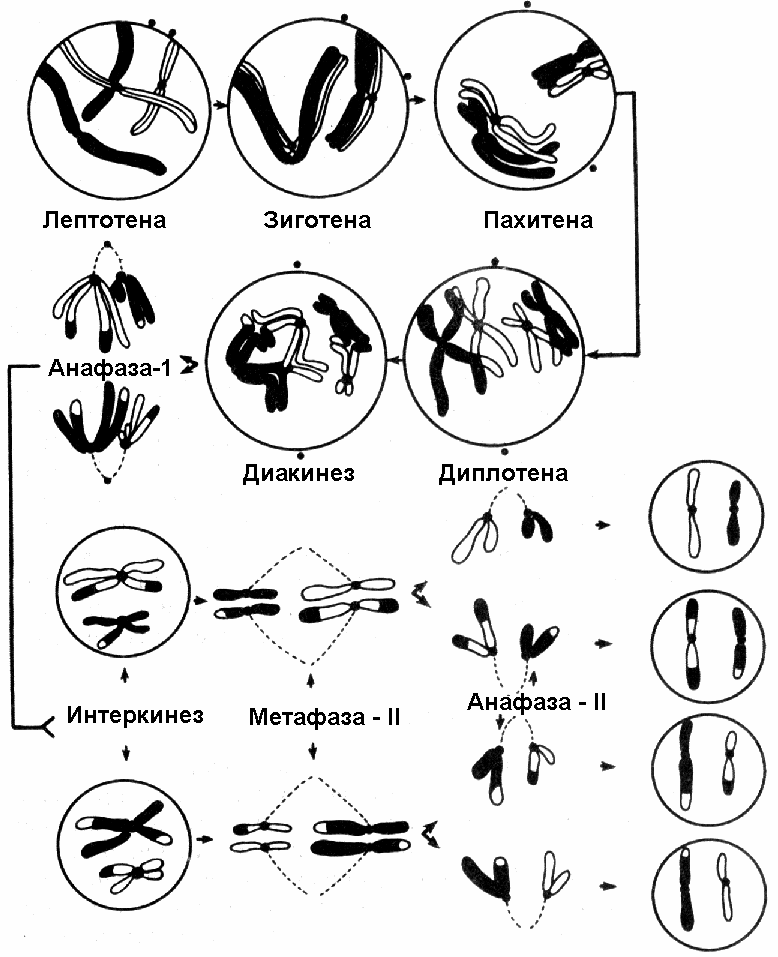
I и II деление мейоза складываются из тех же фаз, что и митоз, но сущность изменений в наследственном аппарате другая (рис. 307). Первое деление мейозаПрофаза 1 (2n; 4с)Самая продолжительная и сложная фаза мейоза. Состоит из ряда последовательных стадий.Лептотена (2n; 4с)Стадия тонких нитей. Хромосомы слабо конденсированы. Они уже двухроматидные, но настолько сближены, что имеют вид длинных одиночных тонких нитей. Теломеры хромосом прикреплены к ядерной мембране с помощью особых структур — прикрепительных дисков.Зиготена (2n; 4с)Стадия сливающихся нитей. Гомологичные хромосомы начинают притягиваться друг к другу сходными участками и конъюгируют. Конъюгацией называют процесс тесного сближения гомологичных хромосом. (Процесс конъюгации также называют синапсисом.)Конъюгация может начинаться в разных точках хромосом (чаще всего с концов, иногда с центромер), а затем распространяться по всей длине. Полагают, что каждый ген приходит в соприкосновение с гомологичным ему геном другой хромосомы. Пару конъюгирующих хромосом называют бивалентом. Биваленты продолжают укорачиваться и утолщаться. Интересен тот факт, что в зиготене гомологичные хромосомы, образуют биваленты, объединяются — четыре хроматиды удерживаются вместе, количество бивалентов равно гаплоидному набору хромосом. Пахитена (2n; 4с)Стадия толстых нитей. Процесс спирализации хромосом продолжается, причем в гомологичных хромосомах он происходит синхронно. Становится хорошо заметно, что хромосомы двухроматидные. Таким образом, каждый бивалент образован четырьмя хроматидами. Поэтому его называют тетрадой. В пахитене наблюдается особенно тесный контакт между хроматидами.Важнейшим событием пахитены является кроссинговер — обмен участками гомологичных хромосом, что приводит к образованию перекреста между несестринскими хроматидами бивалента. В пахитене перекресты еще не видны. Они проявляются позднее в виде хиазм. Кроссинговер приводит к первой во время мейоза рекомбинации генов. Диплотена (2n; 4с)Стадия двойных нитей. Хромосомы в бивалентах перекручиваются и начинают отталкиваться друг от друга. Процесс отталкивания начинается в области центромеры и распространяется по всей длине бивалентов. Однако они все еще остаются связанными друг с другом в некоторых точках. Их называют хиазмы23. Эти точки появляются в местах кроссинговера. В ходе гаметогенеза у человека может образовываться до 50 хиазм.Диакинез (2n; 4с)Хромосомы сильно укорачиваются и утолщаются за счет максимальной спирализации хроматид, а затем отделяются от ядерной оболочки. Происходит сползание хиазм к концам хроматид. Каждый бивалент содержит четыре хроматиды, которые удерживаются друг около друга благодаря наличию хиазм, переместившихся к их концам.В конце профазы I исчезают ядерная оболочка и ядрышко. Биваленты перемещаются в экваториальную плоскость. Центриоли (если они есть) перемещаются к полюсам клетки, и формируется веретено деления. Метафаза I (2n; 4с)Заканчивается формирование веретена деления. Спирализация хромосом максимальна. Биваленты располагаются в плоскости экватора. Причем центромеры гомологичных хромосом обращены к разным полюсам клетки. Расположение бивалентов в экваториальной плоскости равновероятное и случайное, то есть каждая из отцовских и материнских хромосом может быть повернута в сторону того или другого полюса. Это создает предпосылки для второй за время мейоза рекомбинации генов.Центромеры хромосом прикрепляются к нитям веретена деления, но не делятся. Анафаза I (2n; 4с)Нити веретена деления тянут центромеры, соединяющие две хроматиды к полюсам веретена деления. Таким образом, к полюсам расходятся целые хромосомы, а не хроматиды, как при митозе. У каждого полюса оказывается половина хромосомного набора. Причем, пары хромосом расходятся так, как они располагались в плоскости экватора во время метафазы. В результате возникают самые разнообразные сочетания отцовских и материнских хромосом (происходит перемешивание хромосом отца и матери), происходит вторая рекомбинация генетического материала.Телофаза I (1n; 2с)У животных и некоторых растений хроматиды деспирализуются, вокруг них формируется ядерная оболочка. Затем происходит деление цитоплазмы (у животных) или образуется разделяющая клеточная стенка (у растений). У многих растений клетка из анафазы I сразу же переходит в профазу II.Таким образом, в результате первого деления мейоза:
Второе деление мейозаИнтерфаза II (1n; 2с)Характерна только для животных клеток. Кратковременна, репликация ДНК не происходит.Вторая стадия мейоза включает также профазу, метафазу, анафазу и телофазу. Она протекает так же, как обычный митоз. Профаза II (1n; 2с)Хромосомы спирализуются, ядерная мембрана и ядрышки разрушаются, центриоли, если они есть, перемещаются к полюсам клетки, формируется веретено деления.Метафаза II (1n; 2с)Формируются метафазная пластинка и веретено деления: хромосомы располагаются в плоскости экватора, нити веретена деления прикрепляются к центромерам, которые ведут себя как двойные структуры.Анафаза II (2n; 2с)Центромеры хромосом делятся, хроматиды становятся самостоятельными хромосомами, и нити веретена деления растягивают их к полюсам клетки. Число хромосом в клетке становится диплоидным, но на каждом полюсе формируется гаплоидный набор. Поскольку в метафазе 2 хроматиды хромосом располагаются в плоскости экватора случайно по отношению к полюсам клетки, в анафазе происходит третья рекомбинация генетического материала.Телофаза II (1n; 1с)Нити веретена деления исчезают, хромосомы деспирализуются, вокруг них восстанавливается ядерная оболочка, делится цитоплазма.Таким образом, в результате двух последовательных делений мейоза диплоидная клетка дает начало четырем дочерним, генетически различным клеткам с гаплоидным набором хромосом. |