Принципы классификации бактерий. Основные формы и размеры бактерий
 Скачать 254.76 Kb. Скачать 254.76 Kb.
|

|
Липиды и углеводы. В состав вирионов всех сложных(оболочечных) вирусов позвоночных кроме нуклеиновой кислоты и белка входят липиды и углеводы. 50-60% фосфолипиды, 20-30% - холестерин. Липиды обнаруживаются только в суперкапсидной оболочкевирионов и имеют клеточное происхождение. Это связанно с тем, что оболочечные вирусы формируются путем почкования на плазматической мембране клеток. Поэтому суперкапсидная оболочка вирионов представляет собой мембрану клетки-хозяина,модифицированную за счет встроенных в нее вирусных белков – пепломеров. В вирионах в основном обнаруживают фруктозу, сахарозу, галактозу, глюкозамин. Углеводы являются каркасом для локальных участков гликопротеидов, обеспечивают сохранение конформации белковых молекул и защищают от действия протеаз. Компоненты клетки-хозяина. В составе вирионов могут обнаруживаться некоторые компоненты клеток-хозяина. Это могут быть белки или цельные клеточные структуры. Например, в составе некоторых оболочечных вирусов находится белок цитоскелета, рибосомы, клеточные гистоны. Жизненный цикл вирусов. Стратегия вирусного генома. 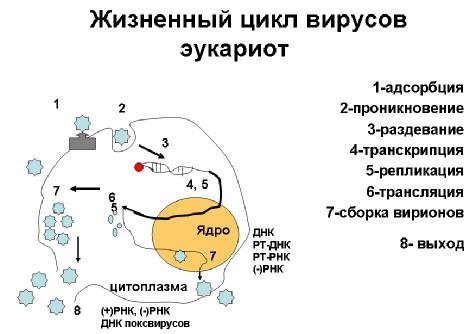 Стратегия генома: Биологической системе, способной к размножению (клетке или вирусу), необходим для жизнедеятельности набор белков , обладающих специфическими функциями (ферментативной, структурной и т.д.). Свойства белка зависят от последовательности аминокислот в полипептидной цепи, а эта последовательность, в свою очередь, определяется последовательностью нуклеотидных звеньев в генетическом материале - ДНК или РНК . Каждому белку соответствует ген , представляющий собой участок нуклеотидной цепи. Совокупность генов, вместилище всего генетического материала клетки или вируса, называют геномом . Схему последовательности событий, которые приводят к реализации генетической информации, т.е. к образованию вирусных белков, а также к репликации носителя генетической информации (ДНК или РНК), называют стратегией генома. В клетке хранилищем генетической информации является двунитевая ДНК, образующая двойную спираль . Для реализации этой информации двойная спираль ДНК должна быть расплетена, после чего на одной из нитей, как на матрице, синтезируется комплементарная ей нить РНК. Эту молекулу РНК, содержащую информацию, необходимую для синтеза белков, называют информационной, или матричной, РНК ( мРНК ). Синтез мРНК на ДНК-матрице называют транскрипцией . Нить мРНК "прочитывается" рибосомой , которая синтезирует белок, причем каждый нуклеотидный триплет является сигналом для включения одной аминокислоты в полипептидную цепь белка. Этот процесс называется трансляцией . Последовательность нуклеотидов в мРНК называют позитивной, или плюс-цепью. Соответственно, та нить ДНК, которая ей комплементарна (и которая используется в качестве матрицы при синтезе мРНК), представляет собой негативную, или минус-цепь. Этот ряд событий, который можно выразить формулой ДНК --> транскрипция --> мРНК --> трансляция --> белок, является одним из двух компонентов стратегии клеточного генома. Он обеспечивает создание всего набора белков, необходимых для жизнедеятельности клетки. Второй элемент клеточной стратегии - репликация ДНК , т.е. воспроизведение клеточного генома. При репликации, как и при транскрипции, нити ДНК расплетаются, но в этом случае, в отличие от транскрипции, на каждой из нитей образуется комплементарная ей нить ДНК, что и приводит к удвоению генома. Этот процесс может быть выражен формулой ДНК--> репликация --> ДНК. Этой простой схемой описывается стратегия любой клетки, но далеко не любого вируса. Даже у ДНК-содержащих вирусов стратегия может быть более сложной, потому что лишь некоторые вирусы содержат, подобно клеточной хромосоме, линейную двунитевую ДНК. Геном других ДНК-содержащих вирусов представлен иными формами ДНК и имеет свои особенности. Стратегия генома РНК-содержащих вирусов еще более резко отличается от клеточной. Для ее характеристики приходится расширить понимание таких терминов, как репликация и транскрипция. Репликацией в этом случае называют воспроизведение геномной РНК независимо от того, используется ли при этом в качестве матрицы РНК или ДНК, а транскрипцией называют синтез только тех молекул вирусной РНК, которые способны функционировать как мРНК, т.е. транслироваться рибосомой с образованием вирусных белков. Цитопатология зараженных клеток. Природа вирусных включений. Значение лабораторной диагностики. ЦИТОПАТОЛОГИЯ ЗАРАЖЕННОЙ ВИРУСОМ КЛЕТКИ Патологические изменения зараженных вирусами клеток обусловлены специфическими и неспецифическими процессами. К неспецифическим процессам относятся процессы, обусловленные изменением проницаемости плазматической мембраны, маргинация хроматина, хромосомные аберрации, пикноз ядер, вакуолизация цитоплазмы. Последнее свойство может приобретать настолько своеобразный и выраженный характер, что превращается в специфический признак некоторых вирусных инфекций. Так, один из вирусов, вызывающий такой процесс,— SV40 — получил название «вакуолизирующий вирус». Специфическими изменениями являются, например, вирусные включения, образование симпластов. Специфические и неспецифические процессы могут привести к деструкции клетки. Цитопатический эффект и его причины.Деструкцию клетки, возникающую при цитолитической инфекции, называют цитопатическим эффектом, а вирус, вызывающий этот эффект, называют цитопатогенным. Большинство вирусов животных являются цитопатогенными, и это свойство лежит в основе патогенеза ряда вирусных инфекций. Цитопатический эффект широко используется в лабораторной диагностике вирусных инфекций для индикации вируса в культуре клеток и выявления антител в сыворотках переболевших. Цитопатический эффект является следствием нескольких причин: 1) нарушение нормальной жизнедеятельности клетки в результате механического повреждающего действия вирусных компонентов на клеточные структуры; 2) повреждение лизосом, в результате чего освобождаются высокоактивные лизосомальные ферменты, вызывающие аутолиз клетки; 3) интенсивное истощение белковых и энергетических ресурсов клетки за счет переключения клеточных ферментов и белок-синтезирующего аппарата на синтез вирусспецифических макромолекул; 4) специфическое повреждающее действие вирусов на клеточные молекулы. Эти причины повреждения клетки различным образом проявляются и сочетаются при разных вирусных инфекциях. Среди РНК-содержащих цитопатогенных вирусов пикорнавирусы оказывают наиболее быстрое и глубокое действие на синтез клеточных белков. Причиной выключения белкового синтеза является блокирование узнавания рибосомой «шапочки» клеточных иРНК. Поскольку РНК вируса полиомиелита транслируется по механизму, независимому от «шапочки», происходит селективное подавление трансляции клеточных иРНК. Вирусные включения.Вирусные включения, выявляющиеся при окрашивании зараженных клеток, являются специфическими морфологическими признаками вирусной инфекции, часто имеющими диагностическое значение. Внутриклеточные вирусные включения были обнаружены гистологами еще в прошлом столетии. Д. И. Ивановский обнаружил в клетках растения, зараженного вирусом табачной мозаики, кристаллоподобное включение, которое впоследствии получило название «кристаллы Ивановского». Позже было доказано, что «кристаллы Ивановского» представляют собой скопление вирусов табачной мозаики. Вирусные включения выявляются в ядре или цитоплазме зараженной клетки. В зависимости от прокрашивания разными красителями включения бывают базофильными и ацидофильными (эозинофильными). Включения при разных вирусных инфекциях различаются по величине, форме, численности. Они могут быть одиночными и множественными, крупными и мелкими, округлыми или неправильной формы. Характерные ядерные включения формируются в клетках, зараженных вирусами герпеса, полиомы, аденовирусами, флавивирусами, вирусом ящура. Характерные цитоплазматические включения формируются в клетках, зараженных вирусами оспы, гриппа, бешенства. Природа включений разнообразна. Большей частью включения представляют собой «вирусные фабрики», т. е. очаги, в которых идет транкрипция и репликация вирусных геномов и сборка вирусных частиц. В клетках, зараженных реовирусом, образуются причудливые серповидные околоядерные включения; при электронно-микроскопическом исследовании они оказались связанными с нитями митотического веретена, в ассоциации с которыми идет репродукция этого вируса. Включения могут представлять собой скопление вирусных частиц, как, например, внутриядерные включения в клетках, зараженных аденовирусами и вирусом полиомы, либо скопление молекул вирусных белков, например ядерные и цитоплазматические включения в клетках, зараженных вируеом гриппа, представляющие собой скопление молекул неструктурного вирусного белка. Некоторые включения содержат только клеточный материал, например ядерные ацидофильные включения в клетках, зараженных вирусами герпеса на поздней стадии инфекции. Бактериофаги, классификация, строение. Бактериофаги [от бактерии, + греч. phagein, поедать] — группа вирусов, паразитирующих в бактериальных клетках. Вирусы, вызывающие гибель инфицированных бактерий, известны как литические бактериофаги. Размножение и выход дочерних популяций вируса из бактерии сопровождается её гибелью и разрушением (лизисом). По морфологии фаги подразделяются на 6 групп: 1) фаги с длинным отростком, чехол которого сокращается; 2) фаги с длинным отростком, чехол которого не сокращается; 3) фаги с короткими отростками; 4) фаги с аналогом отростка; 5)фаги без отростка; 6) нитевидные фаги. По специфичности взаимодействия различают следующие бактериофаги: 1) поливалентные – взаимодействуют с родственными видами бактерий; 2) моновалентные – взаимодействуют с бактериями определенного вида; 3) типовые – взаимодействуют с отдельными типами бактерий данного вида. Строение бактериофагов наиболее полно охарактеризовано на основе изучения Т-фагов кишечной палочки (рис. 5-10). Внешне большинство бактериофагов напоминают сперматозоиды или головастиков Структура типовых бактериофагов. Практическое использование. Они состоят из головки икосаэдрического типа, в которой заключена или ДНК, или РНК. Большинство фагов содержат двунитевую ДНК, замкнутую в кольцо. Хвостовой отросток имеет внутри полый цилиндрический стержень, сообщающийся с головкой, а снаружи – чехол, способный к сокращению наподобие мышцы. Чехол присоединен к воротничку, окружающему стержень около головки. Хвостовой отросток заканчивается шестиугольной базальной пластинкой с шипами от которых отходят нитевидные структуры – фибриллы. Бактериофаги используют: 1) в лабораторной диагностике инфекций при внутривидовой идентификации бактерий, т.е. определения фаговара. 2) фаги применяют также для лечения и профилактики ряда бактериальных инфекций. 3) бактериофаги широко применяют в генной инженерии в качестве векторов для получения рекомбинантных ДНК. Формы взаимодействия бактериофагов с бактериальной клеткой. Взаимодействие фага с бактериальной клеткой. По механизму взаимодействия различают вирулентные и умеренные фаги. Вирулентные фаги, проникнув в бактериальную клетку, автономно репродуцируются в ней и вызывают лизис бактерий. Процесс взаимодействия вирулентного фага с бактерией протекает в виде нескольких стадий и весьма схож с процессом взаимодействия вирусов человека и животных с клеткой хозяина. Однако для фагов, имеющих хвостовой отросток с сокращающимся чехлом, он имеет особенности. Эти фаги адсорбируются на поверхности бактериальной клетки с помощью фибрилл хвостового отростка. В результате активации фагового фермента АТФазы происходит сокращение чехла хвостового отростка и внедрение стержня в клетку. В процессе «прокалывания» клеточной стенки бактерии принимает участие фермент лизоцим, находящийся на конце хвостового отростка. Вслед за этим ДНК фага, содержащаяся в головке, проходит через полость хвостового стержня и активно впрыскивается в цитоплазму клетки. Остальные структурные элементы фага (капсид и отросток) остаются вне клетки. После биосинтеза фаговых компонентов и их самосборки в бактериальной клетке накапливается до 200 новых фаговых частиц. Под действием фагового лизоцима и внутриклеточного осмотического давления происходит разрушение клеточной стенки, выход фагового потомства в окружающую среду и лизис бактерии. Один литический цикл (от момента адсорбции фагов до их выхода из клетки) продолжается 30—40 мин. Процесс бактериофагии проходит несколько циклов, пока не будут лизированы все чувствительные к данному фагу бактерии. Взаимодействие фагов с бактериальной клеткой характеризуется определенной степенью специфичности. По специфичности действия различают поливалентные фаги, способные взаимодействовать с родственными видами бактерий, моновалентные фаги, взаимодействующие с бактериями определенного вида, и типовые фаги, взаимодействующие с отдельными вариантами (типами) данного вида бактерий. Умеренные фаги лизируют не все клетки в популяции, с частью из них они вступают в симбиоз, в результате чего ДНК фага встраивается в хромосому бактерии. В таком случае геномом фага называют профаг. Профаг, ставший частью хромосомы клетки, при ее размножении реплицируется синхронно с геном бактерии, не вызывая ее лизиса, и передается по наследству от клетки к клетке неограниченному числу потомков. Биологическое явление симбиоза микробной клетки с умеренным фагом (профагом) называется лизогенией, а культура бактерий, содержащая профаг, получила название лизогенной. Это название отражает способность профага самопроизвольно или под действием ряда физических и химических факторов исключаться из хромосомы клетки и переходить в цитоплазму, т. е. вести себя как вирулентный фаг, лизирующий бактерии. Лизогенные культуры по своим основным свойствам не отличаются от исходных, но они невосприимчивы к повторному заражению гомологичным или близкородственным фагом и, кроме того, приобретают дополнительные свойства, которые находятся под контролем генов профага. Изменение свойств микроорганизмов под влиянием профага получило название фаговой конверсии. Последняя имеет место у многих видов микроорганизмов и касается различных их свойств: культуральных, биохимических, токсигенных, антигенных, чувствительности к антибиотикам и др. Кроме того, переходя из интегрированного состояния в вирулентную форму, умеренный фаг может захватить часть хромосомы клетки и при лизисе последней переносит эту часть хромосомы в другую клетку. Если микробная клетка станет лизогенной, она приобретает новые свойства. Таким образом, умеренные фаги являются мощным фактором изменчивости микроорганизмов. Структура и химический состав микробной клетки. Особенности метаболизма бактерий. Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядра, называемого нуклеоидом. Имеются дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили. Некоторые бактерии в неблагоприятных условиях способны образовывать споры. Клеточная стенка. В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом толстой клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40-90 % массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos — стенка). В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. На ультратонких срезах бактерий наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид. Функции клеточной стенки: 1. Обусловливает форму клетки. 2. Защищает клетку от механических повреждений извне и выдерживает значительное внутреннее давление. 3. Обладает свойством полупроницаемости, поэтому через нее избирательно проникают из среды питательные вещества 4. Несет на своей поверхности рецепторы для бактериофагов и различных химических веществ. Метод выявления клеточной стенки - электронная микроскопия, плазмолиз. L-формы бактерий, их медицинское значение L-формы - это бактерии, полностью или частично лишенные клеточной стенки (протопласт +/- остаток клеточной стенки), поэтому имеют своеобразную морфологию в виде крупных и мелких сферических клеток. Способны к размножению. Цитоплазматическая мембрана располагается под клеточной стенкой (между ними - периплазматическое пространство). По строению является сложным липид-белковым комплексом, таким же, как у клеток эукариот (универсальная мембрана). Функции цитоплазматической мембраны: 1. Является основным осмотическим и онкотическим барьером. 2. Участвует в энергетическом метаболизме и в активном транспорте питательных веществ в клетку, так как является местом локализации пермеаз и ферментов окислительного фосфорилирования. 3. Участвует в процессах дыхания и деления. 4. Участвует в синтезе компонентов клеточной клетки (пептидогликана). 5. Участвует в выделении из клетки токсинов и ферментов. Цитоплазматическая мембрана выявляется только при электронной микроскопии. |