Принципы классификации бактерий. Основные формы и размеры бактерий
 Скачать 254.76 Kb. Скачать 254.76 Kb.
|

|
Риккетсии и хламидии. Морфология, методы выявления. Особенности биологии. Риккетсии названы в честь американского ученого Риккетса, который описал возбудителя риккетсиоза. Имеют все структуры, присущие прокариотам: клеточную стенку (в ней содержится мурамовая кислота), нуклеоид, рибосомы. Спор, жгутиков, капсул не имеют. Грамотрицательны, окрашиваются по Романовскому-Гимзе в лиловый цвет, по Здродовскому (аналог метода Циль-Нильсена) – в красный. Риккетсии полиморфны, т. е. имеют различные морфологические формы: кокковидные (0,5 мкм); палочковидные (1,5 мкм); бациллярные (2-4 мкм); нитевидные (10-40 мкм). Размножаются риккетсии простым делением, а нитевидные формы – дроблением. Вызывают сыпной тиф идругие риккетсиозы. Хламидии (сhlamydis – плащ). Хламидии выделены в отдельный порядок Chlamydiales, который включает 4 семейства. Ведущие патогенные для человека представители хламидий сосредоточены в семействах Chlamydiaceae иParachlamydiaceae, включающие, соответственно, роды Chlamydia и Chlamydophila. Основными, наиболее важными в патологии человека представителями этих родов являются C. psittaci, C. pneumoniae, C. trachomatis. Хламидии грамотрицательные, очень мелкие (0,5 мкм), сферической формы микроорганизмы с облигатным внутриклеточным паразитизмом. Спор, капсул, жгутиков не образуют. Биологическое своеобразие хламидий состоит в энергозависимом паразитизме и уникальном цикле развития. Имеются 2 стадии жизненного цикла. Одна – инфекционная стадия – элементарные тельца (ЭТ), она приспособлена к внеклеточному существованию; другая – ретикулярные тельца (РТ) – внутриклеточная неинфекционная форма, лабильна, обладает выраженной метаболической активностью. Элементарные тельца имеют размер 0,3 мкм, содержат нуклеоид, в клеточной стенке имеется слой – аналог пептидогликана грамотрицательных бактерий. ЭТ проникают в клетку при фагоцитозе. Из поверхностных мембран клетки хозяина вокруг ЭТ образуется вакуоль и ЭТ превращаются в крупные ретикулярные тельца (диаметр 0,5-1 мкм). Внутри образованной вакуоли РТ многократно делятся. В конечном счете вакуоль через 8-12 циклов деления заполняется этими частицами и превращается в микроколонию (включение). На последней генерации из РТ образуются ЭТ нового поколения. Затем мембрана, которая окружает микроколонию, разрушается, и хламидии выходят в цитоплазму, а далее за пределы клетки. Диагностическое значение имеет обнаружение цитоплазматических включений РТ или мелких ЭТ, которые отличаются от ядра клетки и цитоплазмы по цвету и внутренней структуре. Хламидии вызывают трахому, орнитоз, венерический лимфогранулематоз, бленнорею с включениями. Сравнительная характеристика спирохет и простейших. Методы выявления и классификационное положение. Спирохеты занимают промежуточное положение между бактериями и простейшими. Они имеют черты сходства и различия с теми и другими. С бактериями их сближает спиралевидная форма, недифференцированное ядро. С простейшими их сближает отсутствие клеточной жесткой оболочки, активность и гибкость тела, наличие осевой нити, рецидивирующее течение (в виде приступов) вызываемых ими заболеваний. Методы выявления спирохет (трепонем, боррелий, лептоспир) Оболочка спирохет тонкая, эластичная, содержит большое количество липопротеидов до 70 80% и тонкий фрагментированный слой пептидогликана. Обычными методами трепонемы не окрашиваются, так как при фиксации спиртом оболочка легко разрушается и клетка не в состоянии удерживать красители. Спирохеты так же как и простейшие, плохо окрашиваются: анилиновыми красителями. Для их дифференцирования применяют краску Романовского-Гимзы. Лучше окрашиваются боррелии (в синий цвет), хуже лептоспиры (розовый), еще хуже трепонемы (слабо - розовый). Кроме того, бледная трепонема покрыта мукополисахаридным чехлом, на переднем конце которой под клеточной стенкой имеется скопление протеолитических ферментов, с их помощью она способна активно внедряться в цитоплазму и даже ядро клеток организма хозяина. Бледная трепонема покрыта мукополисахаридным чехлом, на переднем конце бледно трепонемы под клеточной стенкой имеется скопление протеолитических ферментов, с помощью которых она способна активно внедряться в цитоплазму и даже в ядро клеток организма хозяина. Особенности морфологии микоплазм. Принципы классификации и методы выявления. Морфология и размеры микоплазм варьируют в зависимости от возраста культуры, условий и сред культивирования. Микоплазмы полиморфны, но при соблюдении необходимых предосторожностей при фиксации и негативном контрастировании электронная микроскопия позволяет выявлять характерные очертания их клеток. Клетки микоплазм ограничены лишь плазматической мембраной (внутрицитоплазматические мембраны у микоплазм не обнаружены). Отсутствие клеточной стенки у микоплазм определяет их пластичность, которая позволяет им проникать через поры фильтров диаметром 0.22-0.45 мкм. Из-за фильтруемости микоплазмы долгое время путали с вирусами. Сферическая форма клеток характерна для большинства видов микоплазм. Клетки одной и той же микоплазмы могут быть сферической (или несколько вытянутой) формы 0.3-0.8 мкм в диаметре, но могут образовывать длинные (до 100 мкм), иногда ветвящиеся тяжи, которые, проходя фазу коккоидных структур, распадаются на ряд сферических клеток. Коккоидные структуры иногда образуют кольцо. Кольца, содержащие по 4-7 нуклеоидов, впервые наблюдали в бульонной культуре Mycoplasma gallisepticum. Эти превращения являются следствием того, что цитокинез сферических клеток отстает от репликации генома: тяжи и коккоидные структуры мультинуклеоидны. Микоплазмы не образуют покоящихся форм или спор. Подобно другим неспорообразующим бактериям микоплазмы в неблагоприятных условиях переходят в некультивируемое состояние, а также образуют "минимальные тела", не способные к размножению, так как они, вероятно, не содержат ДНК. Некоторые виды микоплазм обладают скользящей подвижностью . Клетки таких микоплазм имеют особые структуры и цитоскелетоподобные образования. Так, клетки M. gallisepticum имеют грушевидную форму; M. pneumoniae также грушевидны, но более вытянуты, а M. mycoides чаще имеют форму тяжей. Методы выявления микоплазм Микоплазмы – это прокариоты малых размеров, имеют только цитоплазматическую мембрану и не способны синтезировать пептидогликан. Это полеморфные микроорганизмы, по форме представляют собой сферические или грушевидные структуры, а также разветвленные или спиральные нити, как правило, неподвижны. Клетки микоплазм, в отличие от других прокариот, не имеют клеточной стенки, окружены ЦПМ. Снаружи ЦПМ находится капсулоподобный слой. Морфологию микоплазм изучают в живом состоянии в фазово-контрастном микроскопе и путем электронной микроскопии ультратонких срезов их клеток. По Граму окрашиваются медленно, грамотрицательны. Вирусы. Природа вирусов. Структура вириона. Вирусы – это облигатные внутриклеточные паразиты, имеющие собственный геном, структурные белки и ферменты, способные репродуцироваться только в чувствительных к ним клетках животных, растений, бактериях. Природа вирусов. Вирусы занимают особое положение по сравнению с другими группами микроорганизмов. Они находятся как бы на грани живой и неживой природы. Природа их в течение всей истории вирусологии была предметом многочисленных исследований и дискуссий. Во внешней среде вирусы, в том числе и фаги, представляют собой совершенно инертные образования, не проявляющие признаков жизни - дыхания, питания, размножения. Многие из них можно превращать в кристаллы. В кристаллическом или в высушенном состоянии их можно сохранять долгие годы. Они ведут себя, как химические соединения. Но стоит им попасть в чувствительные клетки, как у них начинают проявляться все признаки жизни - размножение, наследственность, изменчивость, способность к приспособлению, эволюции. В свободном состоянии вирус - это просто гигантская молекула со всеми особенностями, присущими всем большим молекулам. В живой же клетке он проявляет себя как организм, репродуцирует и мутирует. Так, ВТМ представляет собой то мельчайший организм, то самую крупную молекулу. Некоторые ученые, исходя из способности вирусов к кристаллизации и ферментов к аутокатализу, т. е. катализу под влиянием продуктов, выделяемых в результате самой реакции, относят вирусы к ферментам. Но химический состав вирусов и ферментов отвергает его ферментную природу. Ферменты - белковые вещества, а вирусы кроме белков содержат еще нуклеиновые кислоты, которые направляют синтез белков. Явление кристаллизации зависит от массы и структуры частиц, образующих кристаллическую решетку. Масса бактерий, риккетсий, крупных вирусов слишком велика, чтобы они могли стать структурными единицами кристаллов. Мелкие вирусы, обладающие малыми размерами, могут вследствие действия межмолекулярных сил сцепления и физико-химического строения частиц кристаллизоваться. Различают простые и сложные вирионы. Простые вирионы состоят из нуклеиновой кислоты, окруженной снаружи белковой оболочкой, которую называют капсидом (ящиком, футляром), сложные — имеют дополнительную внешнюю оболочку (суперкапсид, пеплос). Структура и химический состав простых вирионов. В состав простых вирусов, типичным представителем которых является вирус табачной мозаики, входят только капсидные белки, но у некоторых из них содержатся также геномные, или терминальные, ковалентно связанные с концом вирионной нуклеиновой кислоты и участвующие в регуляции вирусного генома. Капсидные белки простоорганизованных вирионов обычно состоят из 1-3 вирусоспецифических белков (полипептидных цепей). При этом каждый из них обладает способностью к самосборке, в начале которой из идентичных полипептидных цепей образуются отдельные структурные элементы (субъединицы) или капсомеры белкового чехла, вслед за чем при их взаимодействии с нуклеиновой кислотой вириона происходит полное самопроизвольное формирование капсида. Количество капсомеров в капсиде вирусов варьирует от трех-шести десятков до многих сотен. У одних вирусов капсомеры имеют овальную или округлую форму, у других - пяти- или шестигранную. По характеру расположения капсомеров вирусы делят на три группы: с кубическим, спиральным и смешанным типом симметрии. Большинство патогенных для человека вирусов имеет кубический тип симметрии. Спиральный тип симметрии нередко встречается среди вирусов растений. Смешанный тип симметрии для простых вирусов нетипичен. Структура и химический состав сложных вирионов. Сложно устроенные вирусы в капсиде имеют много разновидностей белков. Кроме капсидных и геномных белков могут также содержать ферменты, участвующие в репликации и транскрипции вирусного генома, например ДНК- и РНК-полимеразы. Формирование их капсидов и нуклеокапсидов происходит тоже в процессе самопроизвольной сборки, но цикл полного образования сложных вирионов носит многоступенчатый характер. В суперкапсиде сложных вирусов, представляющем собой липидный бислой, в котором расположены пепломеры, превалируют гликопротеиды. Являясь типичными внутримембранными белками, они у большинства вирусов образуют поверхностные «шипы», длина которых достигает 5-10 нм. Чаще всего шипы построены из нескольких молекул идентичного белка, и те вирусы, которые имеют один гликопротеид, естественно, обладают одним типом шипов, при наличии в них двух гликопротеидов - двумя типами. Есть, однако, вирусы, имеющие 2-3 гликопротеида, формирующие один тип шипов. Суперкапсидные вирусные гликопротеиды выполняют две основные функции: 1) распознают специфические клеточные рецепторы и взаимодействуют с ними, что дало повод называть их прикрепительными белками, и 2) обусловливают проникновение вируса в клетки, инициируя слияние его оболочек с клеточными мембранами, вследствие чего их называют белками слияния. Количество углеводов в гликопротеидах может достигать 10% общей массы вириона. Обычными сахарными остатками в них являются сахароза, фруктоза, манноза, галактоза, нейраминовая кислота. Углеводы гликопротеидов обеспечивают сохранение конформации белка и его устойчивость к протеазам. Вирусы. Принципы классификации, морфология вирионов. Вирусы – это облигатные внутриклеточные паразиты, имеющие собственный геном, структурные белки и ферменты, способные репродуцироваться только в чувствительных к ним клетках животных, растений, бактериях. Признаки, используемые для классификации вирусов: тип нуклеиновой кислоты – ДНК или РНК; их структура (однонитевая, двунитевая, линейная, кольцевая, фрагментированная, нефрагментированная с повторяющимися и инвертированными последовательностями); структура, размеры, тип симметрии, число капсомеров; наличие или отсутствие внешней оболочки (суперкапсида); антигенная структура; феномены генетических взаимодействий; круг восприимчивых хозяев; географическое распространение; внутриядерная или цитоплазматическая локализация; чувствительность к эфиру и детергентам; путь передачи инфекции. В 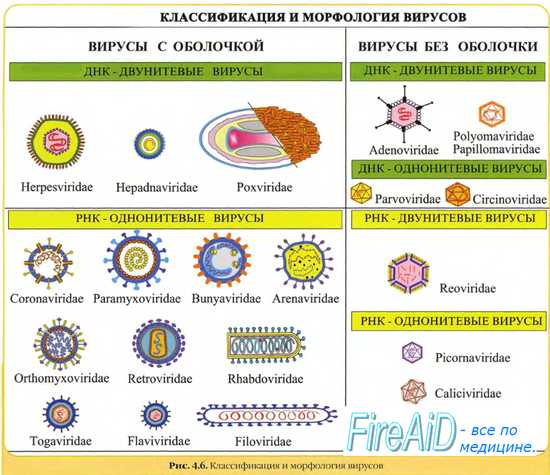 ирусы [от лат. virus, яд] — наименьшие по размерам агенты, имеющие геном, окружённый белковой оболочкой. Вирусы не воспроизводятся самостоятельно, они — облигатные внутриклеточные паразиты, репродуцирующиеся только в живых клетках. Все вирусы существуют в двух формах. В настоящее время известны вирусы бактерий (бактериофаги), грибов, растений и животных. Внеклеточная форма — вирион — включает в себя все составные элементы (капсид, нуклеиновую кислоту, структурные белки, ферменты и др.). Внутриклеточная форма — вирус — может быть представлена лишь одной молекулой нуклеиновой кислоты, так как, попадая в клетку, вирион распадается на составные элементы. Морфология вирусов. Размеры вирусов. Несмотря на внутриклеточный паразитизм, среди вирусов имеются крупные виды, соизмеримые по размерам с микоплазмами и хламидиями. Например, вирус натуральной оспы достигает 400 нм и вполне сравним с риккетсиями (300-500 нм) и хламидиями (300-400 нм). ирусы [от лат. virus, яд] — наименьшие по размерам агенты, имеющие геном, окружённый белковой оболочкой. Вирусы не воспроизводятся самостоятельно, они — облигатные внутриклеточные паразиты, репродуцирующиеся только в живых клетках. Все вирусы существуют в двух формах. В настоящее время известны вирусы бактерий (бактериофаги), грибов, растений и животных. Внеклеточная форма — вирион — включает в себя все составные элементы (капсид, нуклеиновую кислоту, структурные белки, ферменты и др.). Внутриклеточная форма — вирус — может быть представлена лишь одной молекулой нуклеиновой кислоты, так как, попадая в клетку, вирион распадается на составные элементы. Морфология вирусов. Размеры вирусов. Несмотря на внутриклеточный паразитизм, среди вирусов имеются крупные виды, соизмеримые по размерам с микоплазмами и хламидиями. Например, вирус натуральной оспы достигает 400 нм и вполне сравним с риккетсиями (300-500 нм) и хламидиями (300-400 нм). По морфологии выделяют вирусы палочковидные (например, возбудитель лихорадки Эбола), пуле-видные (вирус бешенства), сферические (герпесвирусы), овальные (вирус оспы), а также бактериофаги, имеющие сложную форму (рис. 2-1). При всём разнообразии конфигураций, размеров и функциональных характеристик вирусам присущи некоторые общие признаки. В общем виде зрелая вирусная частица (вирион) состоит из нуклеиновой кислоты, белков и липидов, либо в его состав входят только нуклеиновые кислоты и белки. Химический состав вириона. Простые (безоболочечные) вирусы состоят из нуклеиновой кислоты и белка и представляют собой нуклеопротеиды или нуклеокапсиды. Сложные (оболочечные) вирусы кроме нуклеиновой кислоты и белка содержат также липиды и углеводы. Нуклеиновые кислоты Два типа нуклеиновых кислот – ДНК и РНК. Их структура относительно однообразна: ДНК представляет собой двуспиральную молекулу, РНК – односпиральную молекулу. Функция ДНК заключается в хранении и репликации наследственной информации , таким образом, она является клеточным геномом. РНК представлена в клетке в трех формах: информационной(иРНК), рибосомальной (рРНК), и транспортной (тРНК). Каждая из них выполняет определенную функцию: иРНК образуется в результате транскрипции генома и передает информацию с генома на белок, синтезирующий аппарат клетки, рРНК является структурным компонентом рибосом, тРНК доставляет аминокислоты белок-синтезирующему аппарату клетки. Вотличии от клетки вирусы содержат один тип нуклеиновой кислоты – или ДНК или РНК. Каждая из них выполняет функцию вирусного генома. Структура нуклеиновых кислот у разных вирусов весьма разнообразная. По количеству цепей они бывают одно- и двуспиральными, по форме – линейными и кольцевыми( циркулярными), а также непрерывными и фрагментированными Белки. Белки являются основными компонентами вирионов и состаяляют от 57 до 90% массы вириона. По аминокислотному составу вирусные белки принципиально не отличаются от состава белков животных. В геноме вирусов кодируются две группы белков: структурные, которые входят в состав вирионов потомства, и неструктурные, участвующие в репродукции вируса на разных этапах, но не входящие в состав вирионов. Структурные белки в составе вириона варьируют в широких пределах, что зависит от сложности организации вириона. Простые вирусы 3-4 белка, сложные более 30. Среди структурных белков – капсидные и пепломеры. Капсидные формируют капсид, окружающий нуклеиновую кислоту, геномные белки и ферменты. Пепломеры – белки суперкапсидной оболочки, называемой пеплос. Простые вирусы содержат только капсидные белки, а сложные капсидные и пепломеры. Белки в составе вирусного капсида называются капсомерами, основная функция – защита вирусного генома от неблагоприятных воздействий. Суперкапсидные белки (пепломеры) находятся в липопротеидной оболочке сложных вирусов. Они либо пронизывают липидный бислой вириона, либо не доходят до его внутренней поверхности. Они гликопротеиды, т.е. к молекуле белка в определенных местах прикреплены углеводные цепи. Убольшинства сложных вирусов гликопротеиды формируют на поверхности вириона выступы – шипы длиной 7-10 нм. Неструктурные белки менее изучены.К ним относятся: предшественники вирусных белков, которые существуют в зараженной клетке очень непродолжительное время, а затем нарезаются; ферменты синтеза РНК и ДНК – полимеразы; регуляторы стадий репродукции вирусов; ферменты, модифицирующие вирусные белки – протеиназы и протеинкеназы. |