Принципы классификации бактерий. Основные формы и размеры бактерий
 Скачать 254.76 Kb. Скачать 254.76 Kb.
|

|
Различие в структуре клеточной стенки Гр+ и Гр-бактерий. Химический состав. Функции.
Строение стенки Гр+ бактерий. Структура и функции пептидогликана. Грамположительные бактерии имеют сравнительно просто организованную, но мощную клеточную стенку. Она состоит преимущественно из множества слоев пептидогликана, составляющего до 90% её сухой массы, часто включающих вместо диаминопимелиновой кислоты лизин и уникальные водорастворимые полимеры тейхоевых кислот, состоящих из 8-50 остатков глицерина или рибита, связанных между собой фосфодиэфирными связями. Тейхоевые кислоты [от греч. teichos, стенка] могут составлять до 50% сухого веса клеточной стенки. Известно два типа кислот — рибиттейхоевые (состоят из остатков рибитфосфата и 10-50 остатков спирта) и глицеринтейхоевые (состоят из остатков глицерофосфата и 20 остатков спирта). Клеточная стенка каждого вида содержит только один тип теихоевых кислот (за исключением вида Streptomyces). Тейхоевые кислоты — основные поверхностные Аг многих бактерий. У большей части грамноложительных бактерий также имеются периплазматические теихоевые кислоты, располагающиеся между клеточной стенкой и ЦПМ. Периплазматические кислоты, выявляемые горячей кислотной экстракцией, являются группоспецифичными Аг. Клеточная стенка грамположительных бактерий не содержит ЛПС, но может включать различные белки. Содержание последних весьма вариабельно. Для некоторых бактерий (например, стрептококков группы А) белки служат серовароспецифичными Аг. Опорный каркас клеточной стенки бактерий — пептидогликан (муреин) — гетерополимер, состоящий из повторяющихся дисахаридных групп, соединённых поперечными и боковыми цепочками (рис. 4-3). Остов молекулы пептидогликана — дисахарид. Его образуют N-ацетилглюкозамин и N-ацетилмурамовая кислота, соединённые через р-гликозидные связи. К молекуле N-ацетилмурамовой кислоты присоединяются олигопептиды, образующие боковые цепочки. Строение стенки Гр-бактерий. Функции липополисахарида. Грамотрицателъные бактерии имеют сравнительно тонкую клеточную стенку. В ней выделяют два слоя — пластичный и ригидный. Последний образован одним, редко двумя слоями пептидогликана, содержание которого составляет не более 20% сухой массы клеточной стенки. На пептидогликановом каркасе расположены фосфолипиды, липополисахариды (ЛПС) и белки, образующие пластичный слой. Толщина пластичного слоя значительно превышает размеры монослоя пептидогликана. Его компоненты расположены мозаично и могут образовывать дополнительную внешнюю мембрану либо переходить в капсулу. Липополисахариды клеточной стенки состоят из липидной части (липид А), базисной части молекулы полисахарида (сердцевина) и боковых полисахаридных цепей. Иммуногенные свойства проявляют боковые полисахаридные цепи и сердцевина. Боковые полисахаридные цепи отвечают за антигенную специфичность молекулы липополисахаридов и называются О-Аг. Липидная часть термоустойчива и отвечает за биологические эффекты ЛПС. Структура ЛПС имеет большое диагностическое значение, поскольку разные виды или серовары патогенных грамотрицательных бактерий отличаются друг от друга составом боковых цепей ЛПС внешней мембраны. Цитоплазматическая мембрана, Функциональное значение. 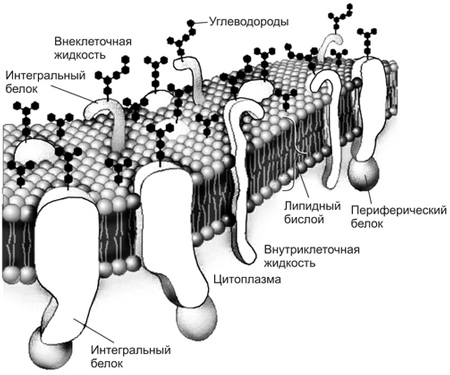 Цитоплазматическая мембрана выполняет многие функции, обеспечивающие жизненные потребности клетки и, в частности, ряд функций необходимых для восприятия и передачи клеткой информационных сигналов. Среди важнейших функций плазматической мембраны можно выделить: отграничение клетки от окружающей се среды с сохранением формы, объема и существенных различий между клеточным содержимым и внеклеточным пространством; перенос веществ внутрь клетки и из нее на основе свойства избирательной проницаемости, активного и других видов транспорта; поддержание трансмембранной разности электрических потенциалов (поляризации мембраны) в покое, ее изменение при различных воздействиях на клетку, генерация и проведение возбуждения; участие в обнаружении (рецепции) сигналов физической природы, сигнальных молекул за счет формирования сенсорных или молекулярных рецепторов и передаче сигналов внутрь клетки; образование межклеточных контактов (плотный, щелевой и десмосомальный контакт) в составе образуемых тканей или при адгезии клеток различных тканей; создание гидрофобного микроокружения для проявления активности ферментов, связанных с мембраной; обеспечение иммунной специфичности клетки за счет наличия в структуре мембраны антигенов белковой или гликопротеиновой природы. Иммунная специфичность имеет значение при объединении клеток в ткань и взаимодействии с клетками, осуществляющими иммунный надзор в организме. Протопласты, сферопласты , L – формы бактерий. Условия их формирования. Протопласты – формы прокариот, полностью лишенные клеточной стенки, образуются обычно у грам «+» бактерий. Сферопласты – бактерии с частично разрушенной клеточной стенкой. Наблюдаются чаще у грам «-» бактерий, реже у грам «+». Образуются в результате разрушения пептидогликанового слоя литическими ферментами: например, лизоцимом или блокирование биосинтеза пептидогликана антибиотиками, например, пенициллином в среде с соответствующим осмотическим давлением. Протопласты и сферопласты имеют сферическую форму или полусферическую, и в 3-10 раз крупнее исходных клеток. В обычных условиях наступает осмотический лизис и они погибают. L-формы бактерий – это фенотипические модификации, или мутанты бактерий, частично или полностью утратившие способность синтезировать пептидогликан клеточной стенки. Свое название они получили в связи с тем, что были выделены и описаны в институте Листера в Англии в1935 г. образуются под воздействием L-трансформирующих агентов – антибиотиков (пенициллина, стрептомицина, полимиксина и др.), аминокислот (глицина, лейцина и др.), фермента лизоцима, ультрафиолетовых и рентгеновых лучей. В отличие от протопластов и сферопластовL-формы обладают относительно высокой жизнеспособностью и выраженной способностью к репродукции.L-формы бактерий полиморфны: встречаются элементарные тельца 0,2-1 мкм, шары- 1-5, большие тела- 5-50, нити. Способностью кL-трансформации обладают практически все виды бактерий.L-формы бактерий в отсутствии фактора, вызвавшего их образование, реверсируют в исходные клетки (для частично утративших синтезировать пептидогликан). L-формам придается большое значение в развитии хронических рецидивирующих инфекций, носительстве возбудителей, длительной персистенции их в организме. Принципы классификации грибов. Классификация грибов довольно сложна и основана главным образом на способах размножения (половое, бесполое) и на структуре вегетативного мицелия. Различают низшие грибы, куда входят два класса: архимицеты и фикомицеты. Высшие грибы объединяют три класса: аскомицеты, базидиомицеты и несовершенные грибы. Все они имеют значение для человека. Фикомицеты широко распространены в природе. К ним относится семействомукоровых грибов — головчатая плесень (рис. 16,2). Клетка мукора состоит из ветвистого неразделенного мицелия, от которого отходят воздушные гифы. Размножаются спорами, которые находятся в мешках-спорангиях на конце плодоносящих гифов (эндоспоры). Половое размножение происходит обычно при недостатке питательных веществ. У человека могут вызывать заболевания — мукорозы, сопровождающиеся лихорадкой, поражением легких и среднего уха. Аскомицеты, или сумчатые грибы, составляют одну из наиболее обширных групп. Размножаются половым путем, образуя в особых сумках — асках определенное число спор. В группу аскомицетов входят как нитевидные грибы (плесени), так и дрожжи. Нитевидные грибы, или плесени, имеют многоклеточный мицелий с межклеточными перегородками в гифах. При вегетативном размножении от одноклеточного гифа-конидиеносца отшнуровываются цепочки спор — конидии (экзоспоры). К плесневым грибам относят два рода: Aspergillus — леечная плесень и Penicillium — кистевик. Аспергиллы, или леечная плесень (рис. 16,3), встречаются на хлебе, фруктах и имеют характерный вид конидиеносца — плодоносящего гифа. От утолщенного конца этого гифа отходят выросты — стеригмы, напоминающие льющиеся из лейки струйки воды. Пенициллы, или гриб-кистевик (рис. 16,4), имеют конидиеносец, который разветвляется на конце, напоминая кисть руки. Плесени широко распространены в природе и играют важную роль в минерализации органических веществ. Некоторые виды плесени используются в промышленности для получения лимонной кислоты, приготовления определенных сортов сыра. Пенициллы являются продуцентами мощного антибиотика — пенициллина. Плесени могут также вызывать заболевания человека. Д 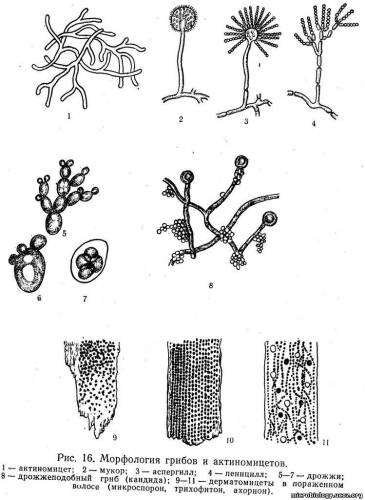 рожжи (рис. 16,5—7)—одноклеточные организмы овальной или удлиненной формы, размером 8— 10 мкм. Внутри клетки имеются ядро, митохондрии, вакуоли, волютин. Дрожжи размножаются почкованием, образуя на поверхности клеток вырост — почку. Процесс почкования длится около 2 ч. За это время в почку переходит часть ДНК, место соединения почки с материнской клеткой утончается и почка отшнуровывается. Спорообразование (половой процесс) наступает после нескольких почкований. Часто споры образуются без предварительного оплодотворения, а иногда и после слияния двух клеток. Внутри сумки — аски образуется 4—8 спор. рожжи (рис. 16,5—7)—одноклеточные организмы овальной или удлиненной формы, размером 8— 10 мкм. Внутри клетки имеются ядро, митохондрии, вакуоли, волютин. Дрожжи размножаются почкованием, образуя на поверхности клеток вырост — почку. Процесс почкования длится около 2 ч. За это время в почку переходит часть ДНК, место соединения почки с материнской клеткой утончается и почка отшнуровывается. Спорообразование (половой процесс) наступает после нескольких почкований. Часто споры образуются без предварительного оплодотворения, а иногда и после слияния двух клеток. Внутри сумки — аски образуется 4—8 спор.Несовершенные грибы (Fungi imperfecti) изучены меньше всего. К ним относят грибы, у которых неизвестен половой способ размножения. Несовершенные грибы .образуют многоклеточный мицелий; размножение осуществляется спорами-конидиями. Многие грибы вызывают заболевания человека, животных и растений. Особенно большое значение имеют возбудители дерматомикозов, вызывающие заболевания кожи, волос и ногтей: трихофитон — возбудитель трихофитии, микроспорон — возбудитель микроспории, ахорион Шенлейна — возбудитель парши, или фавуса (рис. 16,9—11). К несовершенным грибам относят также грибы рода Candida, или дрожжеподобные грибы, которые вызывают у человека, особенно у детей, заболевание слизистой оболочки рта — молочницу (рис. 16,8). Дрожжеподобные грибы имеют округлую форму, размер 8—20 мкм, размножаются почкованием. В отличие от истинных дрожжей им свойственны диморфизм (иногда образуют мицелий) и отсутствие полового размножения. При нерациональном использовании антибиотиков Candida вызывают поражение всего организма — висцеральный кандидамикоз. Морфология грибов. Грибы относятся к царству Fungi (Mycetes, Mycota). Это многоклеточные или одноклеточные нефотосинтезирующие (бесхлорофилльные) эукариотические микроорганизмы с клеточной стенкой. Грибы имеют ядро с ядерной оболочкой, цитоплазму с органеллами, цитоплазматическую мембрану и многослойную, ригидную клеточную стенку, состоящую из нескольких типов полисахаридов, а также белка, липидов и др. Некоторые грибы образуют капсулу. Цитоплазматическая мембрана содержит гликопротеины, фосфолипиды и эргостеролы. Грибы являются грамположительными микробами, вегетативные клетки — некислотоустойчивые. Грибы состоят из длинных тонких нитей (гиф), сплетающихся в грибницу, или мицелий. Гифы низших грибов — фикомицетов — не имеют перегородок. У высших грибов — эумицетов — гифы разделены перегородками; их мицелий многоклеточный. Различают гифальные и дрожжевые формы грибов. Гифальные (плесневые) грибы образуют ветвящиеся тонкие нити (гифы), сплетающиеся в грибницу, или мицелий (плесень). Гифы, врастающие в питательный субстрат, называются вегетативными гифами (отвечают за питание гриба), а растущие над поверхностью субстрата — воздушными или репродуктивными гифами (отвечают за бесполое размножение). Гифы низших грибов не имеют перегородок. Они представлены многоядерными клетками и называются ценоцитными. Гифы высших грибов разделены перегородками, или септами с отверстиями. Дрожжевые грибы (дрожжи), в основном, имеют вид отдельных овальных клеток (одноклеточные грибы). По типу полового размножения они распределены среди высших грибов — аскомицет и базидиомицет. При бесполом размножении дрожжи образуют почки или делятся, что приводит к одноклеточному росту. Могут образовывать псевдогифы и ложный мицелий (псевдомицелий) в виде цепочек удлиненных клеток — «сарделек». Грибы, аналогичные дрожжам, но не имеющие полового способа размножения, называют дрожжеподобными. Они размножаются только бесполым способом — почкованием или делением. Грибы размножаются спорами половым и бесполым способами, а также вегетативным путем (почкование или фрагментация гиф). Грибы, размножающиеся половым и бесполым путем, относятся к совершенным. Несовершенными называют грибы, у которых отсутствует или еще не описан половой путь размножения. Бесполое размножение осуществляется у грибов с помощью эндогенных спор, созревающих внутри круглой структуры — спорангия, и экзогенных спор — конидий, формирующихся на кончиках плодоносящих гиф. Типы грибов. Выделяют 3 типа грибов, имеющих половой способ размножения (так называемые совершенные грибы): зигомицеты (Zygomycota), аскомицеты (Ascomycota) и базидиомицеты (Basidiomycota). Отдельно выделяют условный, формальный тип/группу грибов — дейтеромицеты (Deiteromycota), у которых имеется только бесполый способ размножения (так называемые несовершенные грибы). Сравнительная характеристика актиномицетов и грибов. Классификационное положение, методы выявления. Длительное время актиномицеты считали грибами, однако изучение морфологии и биологических свойств позволило отнести их к бактериям семейства Actinomycetaceae отдела Firmicutes. • В отличие от грибов, актиномицеты не содержат в клеточной стенке хитина или целлюлозы; они не способны к фотосинтезу, а образуемый ими мицелий достаточно примитивен. Также они резистентны к противогрибковым средствам. • С бактериями актиномицеты объединяют отсутствие чётко выраженного ядра, сходство строения клеточной стенки, а также чувствительность к бактериофагам и антибиотикам. Для их роста также оптимальны слабощелочные, но не кислые значения рН среды. Методы выявления и количественного учета актиномицетов. Выделение актиномицетов и их количественный учет проводят методом посева почвенной суспензии на плотные среды — крахмало-аммиачную, крахмало-казеиновую среду, среду Чапека, среду с хитином и др. В качестве селектирующих агентов при выделении актиномицетов из почвы используют ингибиторы, подавляющие рост других микроорганизмов — антибиотики (пенициллин — 1 мг/л, стрептомицин — 25 мг/л, полимиксин — 5 мг/л, нистатин — 50 мг/л, цикло- гексимид — 50 мг/л, пимарицин — 50 мг/л), фенол и другие химические вещества. Выделение актиномицетов методом посева на плотные среды дает возможность в той или иной степени выявить набор видов, содержащихся в той или иной почве, но не дает представления о том, находятся ли актиномицеты в почве в состоянии спор или активно растущего вегетативного мицелия. Для выявления в почве мицелия актиномицетов существует метод прогревания почвенной суспензии при 55— 70°, основанный на том, что гифы большинства актиномицетов гибнут при этой температуре, а споры остаются в жизнеспособном состоянии. Вегетативный мицелий актиномицетов можно также наблюдать при помещении в почву стекол обрастания по Холодному в модификации Рыбалкиной и Кононенко (см. с. 227) и капилляров Перфильева и Габе (см. с. 228). Методы прямого микроскопического учета (метод Виноградского и метод подсчета в люминесцентном микроскопе), применяемые для учета бактерий, могут быть использованы и для учета актиномицетов. Методы исследования почвенных актиномицетов. Культуральные признаки актиномицетов описывают на плотных питательных средах — синтетических и белковых. При описании колоний обычно отмечают цвет воздушного и субстратного мицелия, наличие растворимого пигмента, выделяемого в среду; наличие складчатости колоний (концентрическая или радиальная); поверхность колоний (мучнистая, бархатистая, порошковидная, пушистая). Морфологические свойства актиномицетов — строение колоний и мицелия, его ветвление, строение и расположение спороносцев — можно изучать, просматривая колонии актиномицетов на плотной питательной среде в чашках Петри при малом увеличении микроскопа. Для более детальных исследований получают рост актиномицетов на предметных или покровных стеклах. В агаре на чашках Петри стерильным скальпелем вырезают 1—2 желобка шириной в 1 см во всю глубину агара. Края желобка с помощью петли засевают культурой актиномицета. На засеянные участки желобков накладывают покровные или предметные стекла. Чашки инкубируют обычным способом. При этом актиномицет развивается на поверхности стекла, граничащей со средой. Стекла с развивающейся на них культурой снимают с агара и исследуют прижизненно или после фиксации и окраски. Для знакомства с формой спор актиномицетов делают препарат- отпечаток. Чистое покровное стекло накладывают на газон актиномиц  ета и слегка прижимают к поверхности мицелия. Затем снимают и опускают его в каплю воды на предметном стекле вниз той стороной, на которой имеется налет актиномицет а. Характер поверхности спор выявляют в электронном микроскопе (рис. 42). ета и слегка прижимают к поверхности мицелия. Затем снимают и опускают его в каплю воды на предметном стекле вниз той стороной, на которой имеется налет актиномицет а. Характер поверхности спор выявляют в электронном микроскопе (рис. 42). Для фиксации и окраски мицелия актиномицетов применяют в основном те же методы, что и для бактерий. Для изучения деталей строения спороносцев и спор можно применять фиксацию жаром и окрашивание 0,02%-ным водным раствором кристаллического фиолетового или метиленового синего. |