Рамазанов М. 143. Различие структуры и генетического аппарата прокариот и эукариот
 Скачать 0.9 Mb. Скачать 0.9 Mb.
|

|
2.2 Генетический аппарат прокариот. Наиболее полно изучена структура бактериального генома, в особенности генома E. coli. Основной объем генетической информации бактериальной клетки заключен в ее единственной хромосоме. Размер генома у разных бактерий колеблется от нескольких сотен тысяч пар нуклеотидов (п.н.) до нескольких миллионов п.н. У E. coli он равен 4,6 млн. п.н., а его кодирующая часть составляет 88,6%. В состав бактериальных геномов входят независимые гены и опероны. Работа независимых генов не регулируется другими генами, а их экспрессия носит конститутивный (непрерывный) характер. От соседних генов независимые гены отделены некодирующими участками (спейсерами), которые обычно не транскрибируются. В отличие от независимых генов оперон — это группа рядом расположенных структурных генов, имеющих общую систему регуляции. Обычно эти гены участвуют в осуществлении последовательных этапов какого-либо биохимического процесса. Впервые модель оперона была разработана в 1960 г. французскими биохимиками Ф. Жакобом и Ж. Моно на примере процесса сбраживания лактозы. В систему лактозного оперона входят три структурных гена (Z, Y, A), кодирующие три фермента, участвующие в процессе сбраживания молочного сахара (см. схему). Основным ферментом является β-галактозидаза. К системе регуляции оперона относятся промотор, оператор и ген-регулятор. Промотор расположен перед оператором и является участком, который узнается ферментом РНК-полимеразой, осуществляющим транскрипцию структурных генов. Одна из субъединиц фермента (δ-частица) узнает промотор по специфической последовательности нуклеотидов (блок Прибнова), благодаря чему РНК-полимераза связывается с матрицей.  Фермент начинает транскрипцию, если расположенный рядом с промотором оператор не связан с белком-репрессором, вырабатываемым под контролем гена-регулятора. Отсутствие этой связи обусловлено наличием в клетке субстрата — лактозы, с которой соединяется репрессор. Как только уровень лактозы в клетке падает, регуляторный белок освобождается и садится на оператор, препятствуя тем самым транскрипции структурных генов. Такой тип регуляции носит название негативной индукции, т.к. отсутствие репрессора запускает работу оперона. У прокариот установлены и другие механизмы оперонной регуляции. Например, при синтезе триптофана она может осуществляться по типу репрессии, при котором сам конечный продукт (триптофан) является корепрессором и в комплексе с белком-регулятором, связываясь с оператором, препятствует транскрипции. Объем генома прокариот может увеличиваться, с одной стороны, за счет копирования имеющихся генов, а с другой — за счет включения в геном чужеродной генетической информации. Путями переноса информации у прокариот являются процессы трансформации, конъюгации, трансдукции и транспозиции. Схема процесса трансформации у бактерий 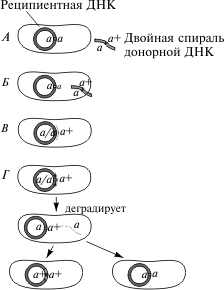 Под трансформацией понимают включение в геном фрагментов чужеродной ДНК, в результате чего клетка приобретает новый признак. Естественную трансформацию наблюдали в смешанных посевах двух штаммов, несущих разные биохимические мутации. О трансформации судили по появлению клеток дикого типа, что возможно только при объединении обеих мутаций в одном геноме и их комплементации. Искусственная трансформация достигается обработкой клеток препаратом ДНК. В обоих случаях клетка, способная воспринимать чужеродную ДНК, находится в особом физиологическом состоянии, которое называется компетенцией. Оно характеризуется увеличением проницаемости клеточной мембраны и активацией ферментативной системы, которая осуществляет перенос фрагмента ДНК через мембрану, разделение его на одиночные цепи и встраивание одиночной цепочки в состав бактериальной хромосомы. Другим каналом для передачи информации у прокариот является процесс конъюгации. Во время конъюгации между двумя бактериальными клетками возникает контакт с образованием цитоплазматического мостика, по которому из одной клетки в другую поступает ДНК.  Основная роль в этом процессе принадлежит половому фактору бактерий — F-плазмиде, внехромосомному носителю информации. Клетки, несущие эту плазмиду (F+), в процессе конъюгации играют роль доноров, а не имеющие ее (F—) — реципиентов. Переход плазмиды из клетки-донора в клетку-реципиент инициирует процесс обмена между ними генетической информацией, т.к. вслед за F-фактором может переноситься бактериальная хромосома. Процесс конъюгации у бактерий гомологичен половому процессу у высших организмов, но отличается от него рядом специфических особенностей. Главная из них состоит в неполной передаче наследственного материала (хромосомы) от донора к реципиенту, благодаря чему образуется частичная зигота — мерозигота, по терминологии Ф. Жакоба и Е. Вольмана. Отсюда весь процесс был назван меромиксисом. В переносе информации от одной бактериальной клетки к другой принимают также участие некоторые бактериальные вирусы — бактериофаги. Это явление получило название трансдукции. Оно было открыто в 1952 г. Дж. Ледербергом и Н. Циндером. Для вирусов, способных переносить информацию, характерен специфический путь развития. Проникнув в клетку, они встраиваются в бактериальную хромосому и могут длительное время находиться в ее составе, уподобляясь ее фрагменту. Это состояние является неактивным, т.к. вирусная ДНК не транскрибируется, и, следовательно, не синтезируются вирусные белки и не образуются новые вирусные частицы. 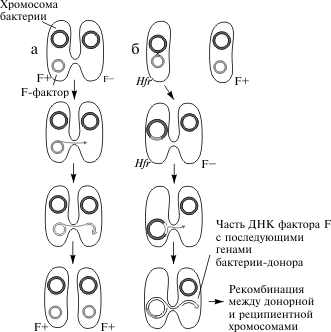 Передача генетического материала в результате конъюгации у E. coli: а — передача F-фактора от донора к реципиенту в скрещивании F+xF–; б — образование линии Hfr в результате интеграции F+ — фактора и передачи бактериальных генов от донорных к реципиентным клеткам в ходе скрещивания F+ x Hfr. Такой путь развития называется лизогенным, а интегрированный вирус — провирусом. Бактериальная клетка, несмотря на присутствие в ней вируса, не подвергается лизису. Однако через какое-то время вирус может активизироваться и выходить из состава хромосомы, “прихватывая” близлежащий фрагмент ДНК бактерии. Следом начинается процесс репликации вирусной ДНК и вместе с нею фрагмента хромосомы. Затем синтезируются вирусные белки и идет сборка новых вирусных частиц, в геном которых включается фрагмент бактериальной ДНК. При этом аналогичный объем собственной информации вирус утрачивает. Клетка в итоге погибает, а освободившиеся из нее вирусные частицы заражают другие клетки, внося в них фрагмент ДНК первого хозяина. Схема процесса трансдукции у E. Coli 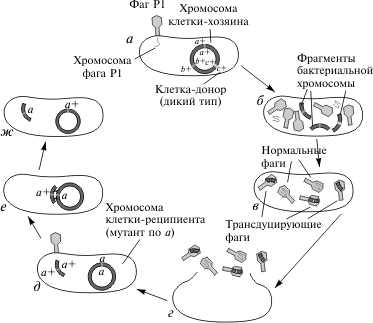 И, наконец, перенос информации в пределах одного генома осуществляется в ходе процесса транспозиции. Транспозиция — это перемещение участка хромосомы из одного места в геноме (локуса) в другой. У бактерий известно два типа транспозирующих элементов: IS-частицы и транспозоны. IS-частицы представляют собой короткие последовательности нуклеотидов, ограниченные концевыми повторами. Они несут информацию о своем перемещении, т.к. в них есть участок, кодирующий структуру фермента транспозазы, осуществляющего вырезание (эксцизию) и встраивание (инсерцию) частицы. Другой информации в IS-частицах нет. В отличие от них транспозоны содержат один или несколько структурных генов, а сложные транспозоны на обоих концах несут еще IS-частицы. Встраивание IS-частиц и транспозонов может вызывать мутации или инактивацию генов, что является одним из возможных путей реорганизации геномов. Схема строения сложного транспозона 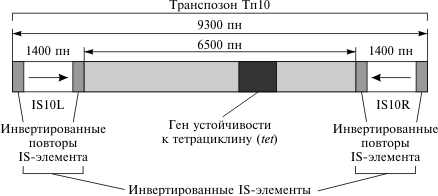 Центральный район, несущий ген или гены сопротивляемости к тетрациклину, фланкирован прямыми или инвертированными IS-элементами. В свою очередь, IS-элементы имеют собственные терминальные инвертированные повторы. В состав бактериального генома входит также плазмидная ДНК. Плазмида — это экстрахромосомный носитель наследственной информации. Количество плазмид в клетке непостоянно. Плазмиды бывают мелкие и крупные, однокопийные и мультикопийные. Однокопийные плазмиды обычно встраиваются в бактериальную хромосому и реплицируются вместе с ней. Их называют эписомами. Мультикопийные плазмиды существуют автономно и реплицируются независимо от бактериальной хромосомы. Число копий их различно. Некоторые плазмиды (например, F-фактор) могут попеременно находиться либо в интегрированном, либо в автономном состоянии. Плазмидная ДНК определяет такие свойства бактериальной клетки, как устойчивость к антибиотикам (R-плазмиды), синтез колицинов — веществ, подавляющих рост других типов бактерий (Col-плазмиды) и др. Многие плазмиды обладают способностью к трансмиссии, т.е. к переходу из одной клетки в другую. Внутри плазмид могут находиться транспозоны. 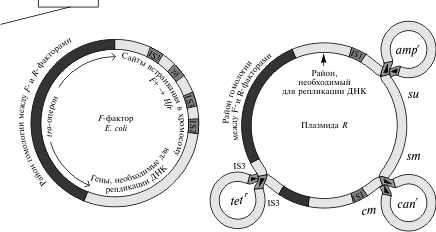 Карты F-фактора и плазмиды R В составе бактериального генома часто обнаруживается вирусная ДНК. Вирусные гены по структуре сходны с бактериальными, но у вирусов есть перекрывающиеся гены. Перекрывание генов происходит в том случае, когда одна и та же последовательность ДНК кодирует структуру двух или трех разных белков за счет изменения рамки считывания. Перекрывающиеся гены были обнаружены Ф. Сенгером в 1977 г. у фага φХ174. Считается, что такая генетическая система является экономичной. Но одновременно мутация в этом локусе может привести к повреждению сразу нескольких генов. 3 Основные различия Важнейшая, основополагающая особенность эукариот связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой. ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки — нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий. В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны. Третье, пожалуй, самое интересное отличие, — это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы — митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом. Ещё одно важное различие между прокариотами и эукариотами — наличие у эукариот эндоцитоза, в том числе у многих групп — фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звёзд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие, среди эукариот появляются первые настоящие, подвижные хищники. Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений — из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать достаточно прочную клеточную стенку из одной молекулы. Это обстоятельство могло заставить их использовать для клеточной стенки иной материал. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе. Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию). Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала. Ещё одно отличие — строение жгутиков. У бактерий жгутиками являются полые нити диаметром 15—20 нм из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотических жгутиков жгутики эукариот изгибаются или извиваются. Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5—10 мкм, когда тот же показатель у эукариот составляет 10—100 мкм. Объём такой клетки в 1000—10 000 раз больше, чем прокариотической. Рибосомы прокариот мелкие (70S-типа). Клетки эукариот содержат как более крупные рибосомы 80S-типа, находящиеся в цитоплазме, так и 70s-рибосомы прокариотного типа, расположенные в митохондриях и пластидах. Видимо, различается и время возникновения этих групп. Первые прокариоты возникли в процессе эволюции около 3,5 млрд лет назад, от них около 1,2 млрд лет назад произошли эукариотические организмы. Заключение Таким образом, мы видим существенные различия как структур, так и генетического аппарата эукариот и прокариот. Эукариоты значительно отличаются более сложной организацией, процессами и составляющими клеток приобретенных в ходе эволюции. В отличие от прокариот эукариоты имеют обособленное ядро, наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной, это митохондрии и пластиды, которые напоминают прокариот. Можно сделать вывод, что эти органоиды являются потомками бактерий, в ходе эволюции вступившие в симбиотическую связь с эукариотами. Существенные различие в размере, к примеру эукариотическая клетка больше прокариотической в 1000-1000 раз. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой. ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки — нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Одним из важнейших отличий эукариот от прокариот это наличие экзонов и интронов. Структурная организация прокариот является более примитивной, нежели организация эукариот. Список литературы
|