Рамазанов М. 143. Различие структуры и генетического аппарата прокариот и эукариот
 Скачать 0.9 Mb. Скачать 0.9 Mb.
|

Базальное тело и механизм его работыБазальное тело представляет собой систему колец, находящихся в плазматической мембране и клеточной стенке бактерий. Два внутренних кольца — M и S-кольца (сейчас чаще рассматриваются как единое MS-кольцо) — являются обязательными элементами, причём M-кольцо находится в ЦПМ, а S — в периплазме грамотрицательных и пептидогликановом слое грамположительных бактерий. Ещё два кольца — P и L — есть только у грамотрицательных бактерий, они расположены в пептидогликановом слое и наружной мембране соответственно, неподвижны и лишь направляют стержень ротора мотора. Вокруг MS-кольца расположены статоры — белковые комплексы MotA4/MotB4, представляющие собой протонный канал (их может быть от 8 до 16). Точный механизм работы базального тела неизвестен. Большинство исследователей полагает, что поступление протона из периплазмы или внешней среды в комплекс MotA4/MotB4 вызывает конформационные изменения белков, благодаря электростатическому взаимодействию или прямому контакту это изменение приводит к поворотуMS-кольца, а его дальнейшее движение возвращает исходную конформацию комплексу и выталкивает протон в цитозоль. У Escherichia coli для одного оборота жгутика требуется перемещение около 1000 протонов. Показано, что жгутик может работать даже у пустых клеточных оболочек при условии, что внешний pH ниже внутреннего. Таким образом, базальное тело преобразует химическую энергию в работу, вращаясь за счёт градиента концентрации протонов или, в редких случаях, ионов натрия(некоторые морские бактерии рода Vibrio, алкалофильные бациллы, Acetobacterium woodii), это вращение осуществляется со скоростью до 100 об/сек, причём его направление может изменяться менее чем за 0,1 сек. Нуклеоид (означает подобный ядру, также известен как ядерная область) — компартмент неправильной формы внутри клетки прокариот, в котором находится генетический материал. ДНК нуклеоида имеет замкнутую кольцевую форму. Такой способ хранения наследственной информации может быть противопоставлен способу эукариот, у которых ДНК упакована в хромосомы и изолирована имеющей мембрану органеллой — ядром. Нуклеоид может быть легко идентифицирован электронной микроскопией при сильном увеличении. Несмотря на то, что он не имеет постоянной формы, он ясно виден на фоне цитоплазмы. При специальном окрашивании нуклеоид также можно различить под световым микроскопом. Нуклеоид состоит в основном из ДНК (примерно 60 %), а также содержит РНК и белки. Последние два компонента представляют собой в основном матричную РНК и белки, регулирующие экспрессию генов бактериального генома. В состав нуклеоида входят также структурные белки, которые способствуют компактизации ДНК, то есть несут функцию, схожую с функцией гистонов в эукариотических клетках. 2 Понятие генетического аппарата. Генетический аппарат - это система взаимодействующих генов в генотипе, который сосредоточен в хромосомном наборе – кариотипе. Генетический аппарат обладает способностью к самовоспроизведению, сохранять постоянной свою организацию, приобретать изменения и воспроизводить их. О. Эйвери доказал, что вещество передающее наследственные признаки – дезоксирибонуклеиновая кислота. ДНК является материальным носителем свойств наследственности и изменчивости, и заключает в себе биологическую информацию – программу развития клетки организма. Современные представления о природе генетического аппарата позволяют выделить три уровня его организации: генный, хромосомный, геномный. Элементарной функциональной единицей наследственности, определяющей возможность развития отдельного признака клетки или организма, является ген. Под признаком понимают единицу морфологической, биохимической, иммунологической, физиологической и любой другой дискретности клетки .или организма в целом, иными словами, отдельное качество или свойство, по которому они отличаются друг от друга. Признак или группа признаков обусловлен действием конкретного гена (с участием или без участия факторов среды). Развитие признака требует синтеза многих веществ, но, прежде всего, - белков, имеющих специфические свойства: ферментативные, структурные, транспортные и т.д. Свойства белковой молекулы определяют ее аминокислотный состав. Он кодируется последовательностью нуклеотидов в ДНК соответствующего 30 гена. Ген – это участок молекулы ДНК, содержащий информацию о первичной структуре одного определенного белка. Гены характеризуются определенными свойствами: Стабильность -способность гена сохранять структуру (ген мутирует редко) Специфичность - каждый ген обладает, присущим только ему, порядком расположения нуклеотидов. Лабильность - способность мутировать или изменяться. Экспрессивность - степень выраженности признака или степень фенотипического проявления данного гена. Пенетрантность - частота проявления гена или его признака. Целостность - ген, кодирующий полипептид (белок) выступает как неделимая частица. Дискретность - это наличие двух субъединиц гена: структурная- это пара комплементарных нуклеотидов, функциональная – это кодон. Плейотропия - множественный эффект действия гена, когда один ген отвечает за проявление нескольких признаков. Аллельность - в генотипе у организмов есть только две формы генов, кодирующих один признак. Амплификация - увеличение количества копий гена. Дозированность - ген обуславливает развитие признаков до определенных пределов дозы. Параллельно дозе усиливается развитие признака Существует несколько вариантов классификации генов. В наиболее распространенном варианте - функциональном - гены делятся на структурные и функциональные. Структурные гены несут информацию о белках, гистонах, а также о последовательностях нуклеотидов в различных видах РНК. Функциональные гены (или последовательности) регулируют работу структурных генов. 1.Структурные гены - гены кодирующие белки, тРНК, рРНК. 2. Регуляторные гены- регулирует функции различных генов. 3. Мигрирующие генетические элементы - транспозоны – перемещаются по геному с помощью ферментов, влияют на активность соседних генов. 4. Псевдогены – копии известных генов лишенные интронов. Они не транскрибируются из-за мутаций в регуляторной области или кодирующей части. 2.1 Генетический аппарат эукариот. Генетический аппарат эукариотической клетки обособлен в виде клеточного ядра, внутри которого располагаются основные носители наследственности — хромосомы. Количество хромосом видоспецифично и колеблется от двух до тысячи. Количество ДНК в клетках эукариот намного выше, чем у бактерий. Ген – это участок молекулы ДНК, содержащий информацию о первичной структуре одного определенного белка. В организме человека насчитывают около 30000 генов. Цистрон – участок ДНК, кодирующий одно полипептидную цепь. Одной из особенностей генома эукариот является структурная и функциональная связь ДНК с белками. Она обусловлена особенностями процесса передачи генетической информации и регуляторной функцией белков. Информация передается от клетки к клетке в процессе сложного процесса клеточного деления (митоза или мейоза). Для полного и точного распределения ее между дочерними клетками в интерфазе происходит процесс удвоения количества ДНК, а в начале деления (профазе) — процесс конденсации интерфазных хромосом. В итоге хромосомы приобретают вид компактных плотных тел. Компактизация хромосом исключает риск их запутывания во время расхождения к разным полюсам в анафазе. В этих структурных преобразованиях хромосом участвуют ядерные белки — гистоны, которые осуществляют суперспирализацию ДНК. Гистоны выступают также в качестве регуляторов матричной активности интерфазных хромосом, т.к. связь гистона с функционирующим участком хромосомы переводит его в гетерохроматическое, т.е. сильно спирализованное и, следовательно, неактивное состояние. Присутствие в составе эукариотических хромосом белков, количество которых удваивается синхронно с удвоением ДНК, делает процесс репликации хромосом более длительным. Характерной особенностью генома эукариот является избыточность ДНК, количество которой намного превышает то, которое необходимо для кодирования структуры всех клеточных белков. Одной из причин избыточности является наличие повторяющихся последовательностей нуклеотидов. Их существование впервые было установлено в конце 60-х гг. ХХ в. американскими исследователями Р. Бриттеном и Д. Девидсоном при изучении кинетики ренатурации ДНК (воссоединения одиночных цепей). В настоящее время установлено, что в составе эукариотической ДНК присутствуют два типа повторов — умеренноповторяющиеся п.н. и высокоповторяющиеся п.н. Умеренные повторы встречаются в виде десятков и сотен копий; средний размер их составляет ≈ 300-400 п.н. Они могут быть прямыми и инвертированными (палиндромы). Между повторами располагаются неповторяющиеся участки ДНК. Высокоповторяющиеся п.н. представляют собой короткие фрагменты ДНК (десятки п.н.), которые представлены большим количеством копий (до 106). В ряде случаев состав оснований в этих повторах отличается от такового в геноме в целом, в результате чего повторы могут образовывать отдельную фракцию с определенной плавучей плотностью. Эта фракция называется сателлитной ДНК. Она никогда не транскрибируется, в связи с чем ее называют также “молчащей”. Установлено, что сателлитная ДНК локализована в гетерохроматических районах хромосом: в теломерах, около центромеры, в ядрышке. Считается, что она выполняет регуляторную функцию, обеспечивая структурные преобразования хромосом во время процесса передачи генетической информации от клетки к клетке. Избыточность ДНК в геноме эукариот в значительной мере создается также за счет того, что в его составе много нуклеотидных последовательностей, которые не кодируют структуру белков. Некоторые из них входят в состав генов, как например, интроны — вставки. Кроме того, есть так называемые сигнальные последовательности, которые не транскрибируются, а служат лишь для связывания белков-регуляторов. К их числу относятся промоторы, участки, контролирующие спирализацию хромосом; участки прикрепления хромосом к веретену и др. Лишь немногие гены присутствуют в эукариотическом геноме в единственной копии. Основная их масса представлена разным числом копий. Расположенные рядом идентичные гены образуют кластеры. Существование кластеров говорит о большой роли дупликаций генов в эволюции геномов. Пример кластеров: гены белков эритроцитов — глобинов. Гемоглобин является тетрамером, состоящим из 4-х полипептидных цепей: 2α и 2β. Каждый тип цепей кодируется генами, организованными в кластер. У человека α-кластер располагается в 11-й хромосоме, а β-кластер — в 16-й хромосоме. β-кластер занимает участок ДНК в 50 тыс. п.н. и включает в себя пять функционально активных генов и один псевдоген. Псевдогены — это нефункционирующие, реликтовые гены, произошедшие в результате мутационных изменений от некогда активных генов. Они не экспрессируются. Гены в составе кластера отделены друг от друга спейсерами — нетранскрибируемыми вставками, в которых иногда могут присутствовать регуляторные участки. Основным отличием эукариотических генов от генов прокариот является то, что большинство из них имеют прерывистую структуру и состоят из кодирующих участков — экзонов и некодирующих вставок — интронов. Длина экзонов от 100 до 600 п.н., а интронов — от нескольких десятков до многих тысяч п.н. Интроны могут составлять до 75% от длины гена. Прерывистая структура генов создает основу для более тонкого контроля их работы. В результате транскрипции прерывистых генов образуется первичный продукт — про-иРНК, которая является полной копией гена и содержит в себе участки, соответствующие как экзонам, так и интронам. В процессе транскрипции участвуют три разных типа РНК-полимераз, которые считывают разные гены. РНКП-I считывает гены, кодирующие структуру разных форм рРНК (5,8S, 18S, 28S). РНКП-II ведет транскрипцию генов, кодирующих структуру белков и некоторых мяРНК. И, наконец, РНКП-III считывает гены 5S рРНК, транспортных РНК и мяРНК. В инициации процесса транскрипции принимает участие белковый комплекс, состоящий из различного числа белковых факторов транскрипции. У млекопитающих в его состав входят 12-14 полипептидов с общей массой в 600 кДА. В регуляции интенсивности транскрипции принимают участие специфические регуляторные участки — энхансеры и сайленсеры. Первые усиливают, вторые ослабляют процесс транскрипции. Они могут быть удалены от промотора на тысячи п.н. Под их контролем синтезируются регуляторные белки. В процессе транскрипции промотор и энхансер (или сайленсер) сближаются за счет структурных изменений ДНК, и регуляторные белки взаимодействуют с факторами транскрипции или с РНК-полимеразой. Для того, чтобы про-иРНК могла играть роль матрицы для синтеза белка, она должна пройти период созревания (процессинг). Главное событие этого периода — удаление из про-иРНК участков, соответствующих интронам, и соединение в единую цепочку оставшихся экзонов. Процесс “сшивания” экзонов называется сплайсингом. В осуществлении сплайсинга большая роль принадлежит малым ядерным РНК (мяРНК) и белкам. Процесс протекает аналогично у всех эукариот. Молекулы мяРНК комплементарно взаимодействуют как с про-иРНК, так и друг с другом. Они обеспечивают удаление интронов и удерживают экзоны вблизи друг от друга. Процесс сплайсинга может носить альтернативный характер, т.е. сшивание экзонов может осуществляться в разных комбинациях. Многие гены содержат десяток и более экзонов, поэтому число вариантов зрелой иРНК = 2n, где n — число экзонов. Альтернативный сплайсинг делает систему записи информации экономичной, так как с одного гена можно считывать информацию для синтеза разных белков. Кроме того, он создает возможность регулирования потока информации в зависимости от потребности клетки в том или ином белковом продукте. Альтернативный сплайсинг, в частности, используется при синтезе иммуноглобулинов, факторов транскрипции и других белков. Полное созревание иРНК включает модификацию обоих ее концов: навешивание кэп-структуры с 5'-конца и присоединение полиадениловой цепочки с 3'-конца. Кэп-структура образуется за счет присоединения к концевому основанию иРНК 5'-конца гуанинового нуклеотида. Схема сплайсинга. 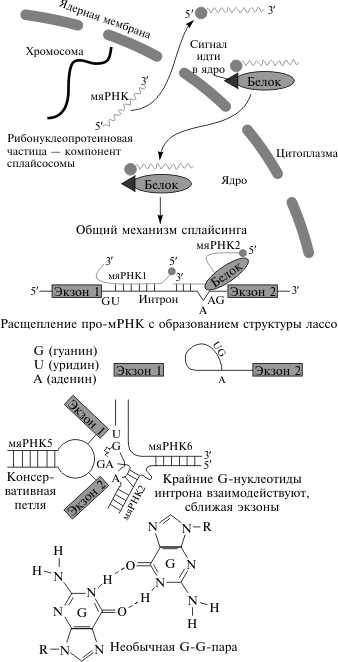 Механизм трансляции у эукариот принципиально не отличается от прокариотического. Однако в обслуживании этого этапа синтеза белка принимает участие значительно большее количество белковых факторов трансляции, чем у бактерий. При характеристике структуры генома эукариот нельзя не сказать о специализированных концевых участках хромосом — теломерах. Теломерная ДНК состоит из многократно повторяющихся коротких блоков нуклеотидов. Впервые теломерная ДНК была изучена у одноклеточных простейших. В ее состав входят блоки по 6-8 пар нуклеотидов. В одной цепи — это блок TTGGGG (G-богатая цепь), в другой — AACCCC (C-богатая цепь). У человека эта последовательность отличается одним основанием TTAGGG, у растений имеется универсальный блок TTTAGGG. Протяженность теломерной ДНК у человека колеблется от 2 до 20 тыс. п.н. Теломерная ДНК никогда не транскрибируется и входит в состав сателлитной ДНК. С теломерными районами хромосом взаимодействует фермент теломераза, который устраняет возникающие в них повреждения. С укорочением теломер в результате потери концевых участков, вызванной снижением активности этого фермента, связывают процесс старения клеток. Существенным отличием функционирования эукариотического генома по сравнению с прокариотическим является многоуровневый характер регуляции действия генов. У прокариот возможен только один тип регуляции — на уровне транскрипции с помощью оперонной системы. У эукариот, благодаря прерывистой структуре генов, к этому типу регуляции добавляется еще посттранскрипционная (сплайсинг, модификация) регуляция и регуляция на уровне трансляции (неоднозначность трансляции). Кроме того, присутствие в хромосомах гистонов позволяет осуществлять групповой контроль за действием генов с помощью механизма структурных преобразований ДНК — перевода участков хромосом из активного (эухроматического) в неактивное (гетерохроматическое) состояние. Такие преобразования иногда затрагивают целые хромосомы и даже весь геном целиком. В качестве примера хромосомного уровня регуляции можно привести образование в клетках женского пола млекопитающих и человека полового хроматина (тельца Барра). Это — крупная гранула хроматина, представляющая собой одну из двух Х-хромосом, максимально конденсированную, и, следовательно, неактивную. Примером инактивации всего генома служит процесс спермиогенеза у животных, во время которого конденсацией охвачены все хромосомы сперматозоида, что делает их неактивными. Это является защитным механизмом для половых клеток в случае повреждения их ДНК (например, при облучении). Возникающие в них мутации, если они не летальны, могут проявиться только при восстановлении функциональной активности мужского генома при дифференциации зародыша. Однако рецессивность большинства мутаций отодвигает их возможное проявление, по крайней мере, до следующего поколения (до перехода в гомозиготное состояние) или вообще исключает его. |