бактериофаги. бактериофагии физ 2. Реферат Бактериофаги
 Скачать 1.35 Mb. Скачать 1.35 Mb.
|

|
Реферат Бактериофаги Белова Ксения 215Б ОМ Содержание Введение Морфология фагов Химический состав фагов Размножение фагов Распространение фагов Проблема бактериофагии на пищевых производствах Заключение Список используемых источников Введение Одним из первых, кто наблюдал и детально описал явление лизиса у бактерий, был один из основоположников отечественной медицинской микробиологии - Н. Ф. Гамалея. В 1896 - 1898 гг. появились его работы, посвященные изучению явления лизиса у сибиреязвенной палочки. Фактор, вызвавший лизис этой бактерии, он назвал бактериолизином. Для развития исследований в области бактериофагии особое значение имели работы французского ученого Д'Эрелля. В 1917 г. он сообщил, что из фекальных масс больных дизентерией ему удалось выделить особый литический фактор (вирус), способный проходить через бактериальные фильтры, размножаться на дизентерийных бактериях и вызывать при этом их лизис. Для обозначения этого вируса Д'Эрелль впервые предложил название бактериофаг. Для обозначения фагов (вирусов микроорганизмов), вызывающих лизис актиномицетов, применяется термин актинофаг, микобактерий - микофаг, кишечной палочки - колифаг, водорослей - цианофаг и т. д. Актуальность данной темы для моей профессии, состоит в том что, на молочных предприятиях одна из главных проблем «явление бактериофагии». Морфология фагов Применение современных электронных микроскопов, а также усовершенствование методов приготовления препаратов для электронной микроскопии позволили более детально изучить тонкую структуру фагов. Оказалось, что она весьма разнообразна и у многих фагов более сложна, чем структура вирусов растений и ряда вирусов человека и животных. 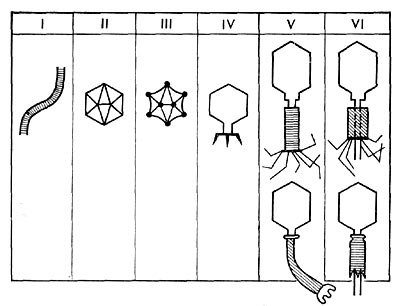 Рис.1 Морфологические типы фагов. Разные фаги отличаются друг от друга не только по форме, величине и сложности своей организации, но и по химическому составу. Оказалось, что фаги, лизирующие микроорганизмы различных групп, могут быть вполне идентичными по своей морфологии. В то же время фаги, активные против одной и той же культуры, могут резко различаться по своей структуре. Так, например, среди фагов, способных лизировать разные штаммы кишечной палочки, выявлены все известные морфологические типы фагов. 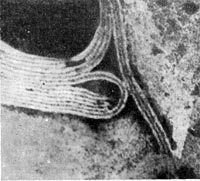 Рис.2 Палочковидные, или нитевидные фаги увел. *400 000 Частицы (или вирионы) большинства известных фагов имеют форму сперматозоида. Они состоят из головки (или капсида) и отростка. Наряду с этим есть фаги, которые состоят из одной головки, без отростка, и фаги, имеющие форму палочки (палочковидные или нитевидные фаги). 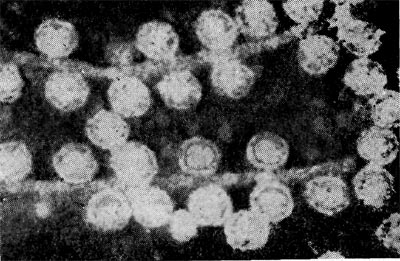 Рис.3 Фаги второго морфологического типа,частица состоит из одной головки увел.*600 000 По форме частиц фаги делятся на шесть основных морфологических типов (групп): палочковидные или нитевидные фаги; фаги, состоящие из одной головки, без отростка; фаги, состоящие из головки, на которой имеется несколько небольших выступов; фаги, состоящие из головки и весьма короткого отростка; фаги, имеющие головку и длинный отросток, чехол которого не может сокращаться; фаги, имеющие головку и длинный отросток, чехол которого может сокращаться. На рис.1 показано схематическое изображение частиц всех шести типов фагов. Размеры фагов принято обозначать в мили-микрометрах или в ангстремах. Фаги первого морфологического типа - палочковидные или нитевидные - выявлены у кишечной, синегнойной, чудесной палочек и других бактерий. Средние размеры их: длина - от 7000 до 8500 А, ширина - от 50 до 80 А (рис.2). Эти фаги отличаются от всех остальных не только большой специфичностью, но и рядом других важных свойств. 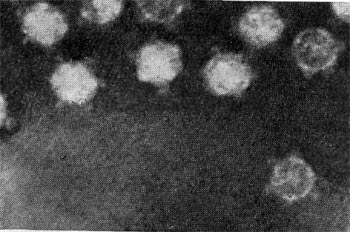 Рис.4 Фаги третьего морфологического типа, от головки отходят небольшие выступы увел. *400 000 Фаги второго морфологического типа. Частица их состоит из одной головки гексагональной (шестигранной) формы на плоскости. Частицы очень мелкие, средний размер их 230-300 А в диаметре (рис.3). У фагов третьего морфологического типа форма и размеры головки такие же, как у фагов второго типа, но у их головок имеются обычно несколько очень коротких выступов (рис.4). Возможно, эти выступы являются аналогами отростков.  Рис.5 Фаг четвертого морфологического типа, частица состоит из головки и короткого отростка увел. *600 000 Фаги второго и третьего морфологических типов отличаются постоянством формы и размеров, независимо от того, против каких микроорганизмов они активны. Эти фаги относятся к мелким формам. Фаги четвертого морфологического типа. Частица состоит из головки, размеры которой варьируют от 400 до 640 А в диаметре, и очень короткого отростка (рис.5). Длина и ширина отростка от 70 до 200 А. Фаги пятого морфологического типа наиболее широко распространены. Головка у частиц гексагональной, формы различных размеров - от 500 до 4250 А в диаметре. Размеры отростка: длина - от 1700 до 5000 А, ширина - от 70 до 120 А (рис.6). Чехол отростка не способен сокращаться. Фаги шестого морфологического типа также широко распространены. Головка частицы различной формы и размеров - от 600 до 1500 А в диаметре, гексагональная. Размеры отростка: длина - от 800 до 2890 А, ширина - от 140 до 370 А. Важной особенностью фагов этой группы является то, что чехол, окружающий отросток, способен сокращаться, в результате чего становится видимым внутренний стержень отростка (рис.7). Головки всех фагов состоят из внутреннего содержимого - нуклеиновой кислоты и окружены белковой оболочкой. Отросток фагов весьма сложен. Он обычно состоит из следующих структур: наружного чехла (или оболочки), внутреннего стержня с канальцем, базальной пластинки, оканчивающейся выступами (типа шипов) и нитевидными структурами. Чехол отростка состоит из субъединиц белковой природы, собранных в спираль. В результате этого он приобретает вид гофрированной трубки. В верхней части отростка многих фагов имеется образование, которое называется воротничком. На рис.8 схематически изображена тонкая структура фаговой частицы. Химический состав фагов Изучение химического состава фагов стало возможно лишь тогда, когда были усовершенствованы методы получения в больших количествах очищенных препаратов фага. В настоящее время изучен химический состав фагов, принадлежащих к разным морфологическим типам и поражающих микроорганизмы почти всех систематических групп. Основными компонентами фагов являются белки и нуклеиновые кислоты. Важно отметить, что фаги, как и другие вирусы, содержат только один тип нуклеиновой кислоты - дезоксирибонуклеиновую (ДНК) или рибонуклеиновую (РНК). Этим свойством вирусы отличаются от микроорганизмов, содержащих в клетках оба типа нуклеиновых кислот. 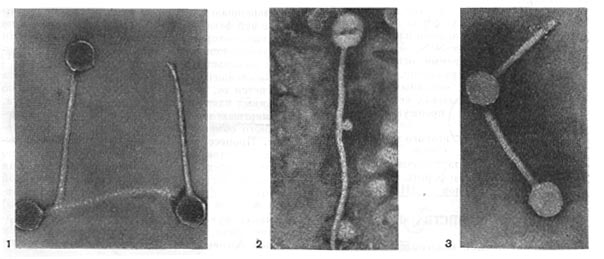 Рис.6. Разные фаги пятого морфологического типа, частица состоит из головки и длинного отростка чехол которого не способен сокращаться. 1,2- увел. X 225 000, 3 - увел. X250 000 Нуклеиновая кислота находится в головке. Внутри головки фагов обнаружено также небольшое количество белка (около 3%). Таким образом, по химическому составу фаги являются нуклеопротеидами. В зависимости от типа своей нуклеиновой кислоты фаги делятся на ДНК-овые и РНК-овые. Количество белка и нуклеиновой кислоты у разных фагов разное. У некоторых фагов содержание их почти одинаковое и каждый из этих компонентов составляет около 50%. У других фагов соотношение между этими основными компонентами может быть различно. 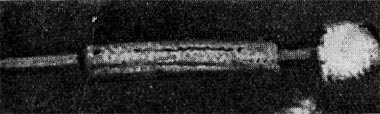 Рис.7. Фаг шестого морфологического типа, частица состоит из головки и длинного отростка, чехол которого способен к сокращению. Увел, около 400 000. Кроме указанных основных компонентов, фаги содержат в небольших количествах углеводы и некоторые преимущественно нейтральные жиры. 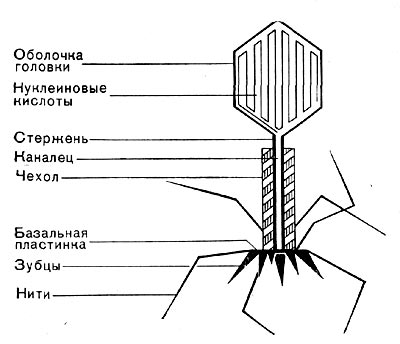 Рис.8. Схема строения фаговой частицы. Все известные фаги второго морфологического типа РНК-овые. Среди фагов третьего морфологического типа встречаются как РНК-овые, так и ДНК-овые формы. Фаги остальных морфологических типов - ДНК-овые. Размножение фагов Взаимоотношения между фагом и чувствительной к нему клеткой очень сложны и не всегда завершаются лизисом клетки и размножением в ней фага. Мы рассмотрим такую инфекцию клетки, которая заканчивается гибелью клетки и размножением в ней фага. Такая инфекция называется продуктивной. Важнейшей особенностью размножения фага является то, что оно может происходить только в живых клетках, находящихся в стадии роста. В мертвых клетках, а также продуктах клеточного обмена размножение фага не происходит. Процесс размножения фага весьма сложный и состоит из следующих последовательно протекающих этапов (рис. 9): 1) адсорбция фаговой частицы на поверхности микробной клетки; 2) проникновение содержимого головки фаговой частицы (нуклеиновой кислоты) в микробную клетку; 3) внутриклеточное развитие фага, заканчивающееся образованием новых фаговых частиц; 4) лизис клетки и выход из нее новых фагов. Время с момента инфицирования клетки фагом до лизиса клетки называется латентным или скрытым периодом. Продолжительность этого периода различна для разных типов фага, зависит от окружающей температуры, состава среды и других факторов. Латентный период фагов, специфичных для одних бактерий, 15-40 мин, для других - 5 ч и более. У фагов актиномицетов латентный период может быть еще продолжительнее. При низкой температуре латентный период значительно увеличивается. 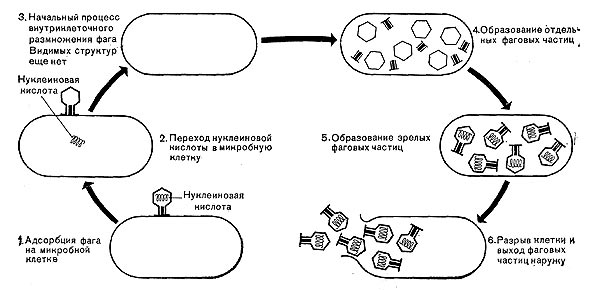 Рис.9. Схема размножения фага. Из всех этапов размножения фага наиболее изучен первый - адсорбция. Адсорбция фага на клетке - реакция весьма специфичная. В клеточной стенке бактерий имеются особые структуры (рецепторы), к которым могут прикрепиться фаги. Адсорбируются на рецепторах только те фаги, к которым чувствительна клетка. Фаги, имеющие отростки, прикрепляются к микробной стенке свободным концом отростка. Нитевидные фаги, а также фаги, не имеющие отростков, адсорбируются не на микробной стенке, а на нитевидных структурах, окружающих стенку, - фимбриях. Описаны фаги, которые прикрепляются отростком к бактериальным жгутикам. У некоторых фагов процесс адсорбции может осуществляться лишь в том случае, когда в среде имеются определенные вещества - кофакторы: аминокислоты (триптофан, тирозин и др.) или соли (кальциевые, магниевые). На конце фагового отростка имеется особый фермент типа лизоцима. После адсорбции фага под влиянием этого фермента происходит растворение стенки микробной клетки и содержимое головки фага - нуклеиновая кислота - перекачивается в микробную клетку. Этим завершается второй этап процесса размножения фага. Остальные структуры фаговой частицы - оболочка головки, отросток и его субструктуры - внутрь инфицированной фагом клетки не попадают. Их роль заключается в обеспечении сохранности фаговой частицы, находящейся вне клетки, и содействии проникновению фаговой нуклеиновой кислоты в клетку при инфекции. 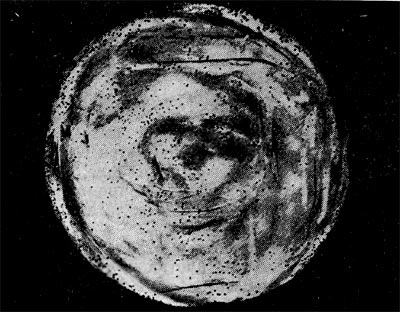 Рис.10. Мелкие (едва видимые) негативные колонии актинофага. Увел. 6 : 10. У нитевидных фагов, в отличие от других видов фагов, внутрь клетки проникает весь белок или его часть. После проникновения нуклеиновой кислоты фага в клетку начинается сложный процесс внутриклеточного размножения фага. Под влиянием нуклеиновой кислоты фага резко изменяется весь обмен микробной клетки. Основные процессы, протекающие в инфицированной клетке, направлены на образование новых фаговых частиц. Вначале формируются отдельно головки и отростки, которые затем объединяются в зрелые фаговые частицы. К этому времени внутри клетки образуется особый литический фермент, который вызывает лизис клетки изнутри. Клетка распадается, и новые зрелые частицы фага выходят наружу 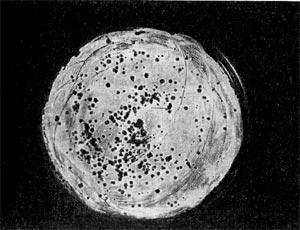 Рис. 11 Мелкие негативные колонии актинофага Увел. 8 : 9. Количество новых фаговых частиц, образуемых одной клеткой при фаговой инфекции, называют выходом фага или его урожайностью. Выход фага зависит от свойств данного фага и не зависит от клетки-хозяина и ее размеров. Одни фаги отличаются очень низким выходом (5-50 частиц на клетку), у других выход значительно выше (от 1000 до 2500). Особенно высоким выходом отличаются мелкие РНК-овые фаги (свыше 20 000 частиц на клетку). Если большое количество бактериальных клеток смешать с небольшим количеством фаговых частиц, то процесс размножения фагов проходит несколько циклов. Вначале инфицируется часть клеток. Первое потомство фага инфицирует оставшиеся клетки - происходит второй цикл, за ним может следовать третий и т. д., пока не будут лизированы все чувствительные к данному фагу клетки. Среди фагов встречаются такие, размножение которых возможно лишь при наличии в среде определенных кофакторов. Одни из этих веществ, как уже указывалось, необходимы для адсорбции фага; другие - для внутриклеточного размножения фага. 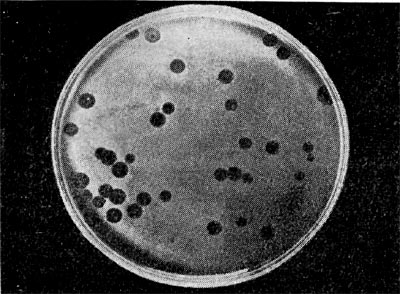 Рис.12. Негативные колонии бактериофага средней величины. Увел. 1:1. Большинство фагов вызывают при размножении лизис клетки и ее гибель. Лишь в последнее время было установлено, что при инфицировании клетки нитевидными фагами возможно размножение их без гибели клетки. Если произвести рассев по поверхности агаризованной питательной среды в чашках Петри смеси фага и чувствительных к нему микробов и чашки выдержать в термостате, то происходит лизис клеток в результате размножения фага. Если взять большое количество частиц фага, то лизируется большая часть или весь выросший газон культуры. Если количество фаговых частиц таково, что они распределяются только на отдельных участках газона, лизируя в этих местах культуру, то возникает колония фага. Эти колонии фага получили название бляшек, стерильных пятен. Правильнее их называть негативными колониями. Каждая негативная колония состоит из десятков и сотен миллионов фаговых частиц. Размер негативных колоний и их форма зависят в первую очередь от свойств фага, а также от состава среды и культуры микробов. У одних фагов негативные колонии очень мелкие и еле видимы невооруженным глазом, другие достигают 10 мм в диаметре и более. Колонии бывают светлые и четкие, когда лизировалась вся культура, или мутноватые, когда лизировались не все клетки. Вокруг негативных колоний некоторых фагов могут возникнуть различной формы и величины ореолы. На рисунках 10-14 показаны негативные колонии разных фагов. Морфология негативных колоний служит одним из признаков, которым пользуются при дифференциации фагов. Распространение фагов В настоящее время найдены, фаги, лизирующие клетки микроорганизмов, принадлежащих ко всем систематическим группам, как патогенных для человека, животных и растений, так и сапрофитных (непатогенных). До недавнего времени не было ясно, существуют ли фаги против плесневых грибов и дрожжей. В последние годы найдены фаги, активные против грибов родов пенициллов, аспергиллов и других, а также против некоторых дрожжей. Интересно отметить, что вирус удалось выявить и у тех видов пенициллов, которые применяются в промышленности для получения пенициллина. Не выявлены вирусы, активные против простейших животных, а также истинных спирохет.  Рис.13. Крупные негативные колонии актинофага. Увел. 1:1. В природных условиях фаги встречаются в тех местах, где есть чувствительные к ним бактерии. Чем богаче тот или иной субстрат (почва, вода, выделения человека и животных и т. д.) микроорганизмами, тем в большем количестве в нем встречаются соответствующие фаги. Так, фаги, лизирующие клетки всех видов почвенных микроорганизмов, находятся в почвах. Особенно богаты фагами черноземы и почвы, в которые вносились органические удобрения. Фаги, активные против разных видов кишечной, дизентерийной, тифозной и паратифозной палочек, часто встречаются в содержимом кишечника человека и животных, сточных водах и загрязненных водоемах. Фаги фитопатогенных микроорганизмов успешнее всего выделяются из остатков растений, пораженных этими микробами. 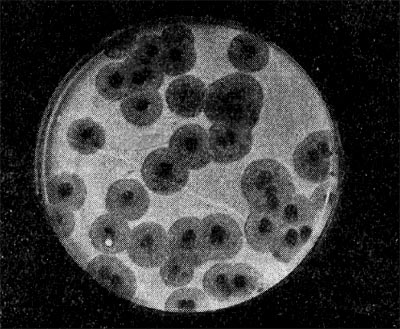 Рис.14. Негативные колонии актинофага, окруженные ореолом угнетенного роста тест-культуры. Увел. 10 : 8. Фаги молочнокислых стрептококков в большом количестве встречаются в молочных продуктах. Итак, те субстраты, на которых развиваются определенные формы микроорганизмов, также благоприятны для существования соответствующих фагов. Проблема бактериофагии на пищевых производствах Широкому распространению фагов молочнокислых бактерий способствует то, что они являются лизогенными, т. е. содержат внутри клеток фаги. В результате этого в молоке сразу же после доения уже есть фаги. Молоко, поступающее на молочные заводы, часто содержит фаги в значительном количестве (1 млн. и более частиц в 1 мл молока). Следует отметить, что все фаги, даже активные против патогенных микробов, совершенно безвредны для людей и животных. Современное производство сыра основано на применении пастеризованного молока. Однако пастеризация не убивает всех имеющихся в молоке фагов. Охлажденное после пастеризации молоко разливают в специальные ванны, в которые вносится закваска, состоящая из чистых культур молочнокислых стрептококков. Стрептококки вызывают сквашивание молока. Получаемый сгусток молока подвергается дальнейшей переработке. Работники сыродельных заводов давно обратили внимание на то, что в ряде случаев активность молочнокислых микробов закваски резко снижается, что приводит к плохому сбраживанию молока. Это явление может быть вызвано разными причинами. Но чаще всего оно вызывается фагами, которые лизируют полностью или частично культуры заквасок. В результате этого процесс молочнокислого брожения полностью останавливается или интенсивность его резко снижается. На сыродельных заводах, как правило, применяют закваски, состоящие не из одной культуры, а из смеси различных культур молочнокислых стрептококков. При применении смешанной закваски под влиянием фага лизируется одна или две культуры, другие же продолжают процесс молочнокислого брожения. Заключение Явление бактериофагии фактически превратилось в самостоятельную область биологии со своими специфическими разделами. Бактериофаги применяют в качестве индикаторов загрязнения воды патогенными энтеровирусами. Бактериофаги по своим биологическим свойствам стоят к энтеровирусам ближе, чем бактерии группы кишечных палочек или другие санитарно-показательные микроорганизмы. Новейшая медицина предлагает лекарства на основе бактериофагов. Фаги могут стать более эффективными, чем обычные антибиотики. Белки, являющиеся «действующим веществом» вируса, по сути, являются прообразами возможных лекарств нового поколения, к которым бактерии не смогут выработать устойчивость. Ученым удалось установить структуру белка, способного блокировать туберкулезную палочку. Список использованных источников Микробиология молока и молочных продуктов: Учебник для вузов. П.П.Степаненко– Сергиев Посад: ООО «Все для вас-Подмосковье», 1999, 415с. Жизнь растений. Том 1. Введение Бактерии и актиномицеты. Н.А. Красильников, А.А. Уранов М. «Просвещение», 1974, 487с. http://www.gazeta.ru/science/2010/01/19_a_3313540.shtml Сыроделие: технологические, биологические и физико-химические аспекты. Под редакцией С.А. Гудкова. - М.: ДеЛи, 2003. - 800 с Методы выделения, изучения и культивирования микроорганизмов: Учебник для вузов. Т. И. Громовых, КрасГУ, 2006. - 159 с |