готовая генетика. Решение задач. 19 Выпишите и дайте объяснение всем терминам, встречавшимся вам при выполнении работы. 21 Список литературы. 25
 Скачать 57.65 Kb. Скачать 57.65 Kb.
|

Содержание.Основные этапы развития генетики. 3 Понятие о наследственности и изменчивости. 13 Мутации. 15 Возвратное и анализирующее скрещивание. 17 Решение задач. 19 Выпишите и дайте объяснение всем терминам, встречавшимся вам при выполнении работы. 21 Список литературы. 25 Основные этапы развития генетики.С поразительной быстротой открытия Г. Де Фриза, К. Корренса и Э. Чермака были подхвачены другими учеными, положив начало бурному развитию генетических исследований. Это показывает, что для понимания открытия Г. Менделя решающей была общая подготовленность биологов. В этом плане нельзя не упомянуть работы известного английского ученого У. Бэтсона (1861–1926), который высказал ряд пророческих мыслей о необходимости изучения поведения отдельных признаков и статистического учета всех появляющихся в потомстве типов. Подобные мысли высказывал и Г. Де Фриз, а также английский ученый К. Пирсон, который разработал статистические методы для биологии, положив начало развитию биометрии. В течение всей последующей истории генетика прошла ряд периодов, каждый из которых характеризовался теми или иными открытиями. Эти периоды тесно связаны между собой и условно их можно разделить на 6 этапов. Первый этап (с 1900 до 1912 г.) – это период триумфального развития менделизма, подтверждение законов Г. Менделя на различных культурах (лабораторных грызунах, курах, бабочках и др.), в результате чего выяснилось, что законы наследственности носят универсальный характер. Особенно примечательными являются работы У. Бэтсона по анализу наследования формы гребня у кур (при скрещивании кур с розовидной и гороховидной формой наблюдалось появление новой ореховидной формы). Выдающееся значение имели работы А. Гаррода, опубликованные в 1902 и 1908 гг. по «врожденным нарушениям метаболизма» у человека. К сожалению, значение работ этого ученого как первых работ в области биохимической генетики, стало понятным только в наши дни. Уже в 1909 г. У. Бэтсон опубликовал сводку, где было перечислено более сотни признаков растений и столько же видов животных, для которых было доказано наследование признаков согласно законам Г. Менделя. В 1906 г. английский ученый Р. Пеннет предложил использовать «решетку Пеннета» для определения генотипов потомства при скрещивании. Сложились такие генетические понятия, как гомозигота и гетерозигота, аллеломорф (термины введены У. Бэтсоном в 1902 г.), ген, фенотип, генотип (термины предложены датским генетиком в 1909 г.). Термин аллеломорф был заменен В. Иоганнсеном на аллель в 1926 г. Его определение звучит следующим образом: аллели – формы состояния гена, вызывающие фенотипические различия, но локализованные на гомологичных участках гомологичных хромосом. В 1906 г. У. Бэтсон сформулировал «правило чистоты гамет». Наряду с наиболее характерными открытиями для этого периода развития менделевской генетики, в эти годы зародились некоторые новые важные направления, получившие свое развитие в последующие годы. Уже в 1902 г. два ученых Т. Бовери в Германии и У. Сэттон в США одновременно обратили внимание на параллелизм в поведении хромосом при мейозе и оплодотворении с передачей признаков потомству, что послужило предпосылкой для создания хромосомной теории наследственности. В 1901 – немецкий ботаник К. Корренс высказал предположение о том, что гены локализованы в хромосомах в линейном порядке, а в 1903 г. эту же мысль подтвердили американский ученый У. Сэттон и немецкий ученый Т. Бовери. Началось изучение внезапно возникающих и стойко наследующихся изменений наследственного материала – мутаций. Г. Де Фриз в 1901 г. сформулировал мутационную теорию, во многом совпавшую с теорией гетерогенеза (1899) русского ботаника С. И. Коржинского. Согласно мутационной теории наследственные признаки не являются абсолютно константными и могут скачкообразно изменяться, вследствие мутирования их задатков. Мутационная теория Г. Де Фриза сыграла большую роль для развития представлений об изменчивости организмов. Ошибочными были лишь его суждения, что мутации в организме возникают самопроизвольно, под влиянием каких-то чисто внутренних причин и не зависят от внешних воздействий. В 1906 г. английские ученые У. Бэтсон и Р. Пеннет в опытах с душистым горошком обнаружили явление сцепления наследственных признаков, а другой английский генетик Л. Донкастер в опытах с бабочкой крыжовенной пяденицей открыл сцепленное с полом наследование. Большое значение для зарождения популяционной генетики сыграли работы, опубликованные в 1908 г. английским математиком Г. Харди (1877–1947), который вывел формулу распределения генотипов в свободно скрещивающихся популяциях, и немецким врачом В. Вайнбергом (1862–1937), который независимо от Г. Харди установил такую же закономерность в отношении человеческой популяции. Поэтому ныне принято именовать установленную ими закономерность как «закон Харди-Вайнберга». На этом этапе истории генетики была принята и получила дальнейшее развитие менделевская, по существу умозрительная концепция гена как материальной единицы наследственности, ответственной за передачу отдельных признаков в ряду поколений организмов. Второй этап (с 1912 до 1925 г.) – это этап создания и развития хромосомной теории наследственности. Ведущую роль в этом сыграли американские ученые Т. Морган (1861–1945) и три его ученика А. Стертевант (1891–1970), К. Бриджес (1889–1938) и Г. Меллер (1890– 1967). Блестящие работы этих ученых в период в 1910 по 1925 г. показали, что наследственные задатки – гены – лежат в хромосомах и что передача признаков определяется судьбой хромосом при созревании половых клеток и оплодотворении. Работы Т. Моргана помогли значительно глубже проникнуть в тонкое строение клеточного ядра и показали возможность построения карт хромосом с указанием точного расположения на них генов (первую карту построил в 1913 г. А. Стертевант совместно с Т. Морганом для Х-хромосомы дрозофилы). В 1911– 1912 гг. Т. Морган разработал хромосомную теорию наследственности, которая прочно опиралась не только на генетические данные, но и на наблюдениях за поведением хромосом в митозе и мейозе, на роли ядра в наследственности. В 1916 г. американский генетик Г. Меллер сформулировал теорию линейного расположения генов на хромосоме. В 1916 г. американский биолог К. Бриджес сформулировал балансовую теорию определения пола. Установил связь между группами сцепления и хромосомами. Значение работ Т. Моргана и его сотрудников для развития генетики огромно. Хромосомная теория наследственности была крупнейшим после Г. Менделя достижением биологии, а проблема гена, поднятая Т. Морганом, стала центральным вопросом генетики, не потерявшим актуальности и сегодня. Все дальнейшее развитие генетики и других биологических дисциплин (цитологии, эмбриологии, биохимии, эволюционного учения и др.) шло в свете этой теории, а на современном этапе послужило основой зарождения молекулярной биологии. В этот же период времени стали развиваться важные для практики направления генетики. К ним можно отнести, начатые еще в 1908 г. шведским генетиком Н. Нильсоном-Эле (1873–1949), работы по изучению закономерностей наследования количественных признаков на примере окраски зерен овса и пшеницы. Получили развитие работы по изучению механизмов гетерозиса (работы американских генетиков Э. Иста и У. Джонса). Благодаря работам Н. И. Вавилова, начала развиваться частная генетика культурных растений. Все это имело большое значение для развития селекционной науки и внедрения в селекционную практику эффективных приемов, опирающихся на достижения генетики В период с 1912 г. по 1925 г. происходило становление отечественной генетики. Уже в первые послеоктябрьские годы сформировался ряд известных школ, возглавляемые крупными учеными – Н. К. Кольцовым (1872– 1940), Ю. А. Филипченко (1882–1930) и Н. И. Вавиловым (1887–1943). Н. К. Кольцов создал Институт экспериментальной биологии, из стен которого вышли известные генетики Б. Л. Астауров, Е. И. Балкашина, Н. К. Беляев, А. Е. Гайсинович, С. М. Гершензон, Я. Л. Глембоцкий, Н. П. Дубинин, В. С. Кирпичников, А. А. Малиновский, И. Б. Паншин, И. А. Рапопорт, П. Ф. Рокицкий, Д. Д. Ромашев, В. В. Сахаров, А. С. Серебровский, Н. В. Тимофеев-Ресовский, В. П. Эфроимсон и др. Все они в той или иной степени восприняли методологию своего учителя. Н. К. Кольцов внес большой вклад в развитие генетики. Он первым разработал гипотезу о молекулярном строении и матричной репродукции хромосом, предвосхитив этим более поздние открытия молекулярной биологии. Выдвинул идею о связи гена с определенным химическим веществом (к сожалению, ошибочную, указав на белок), создал первую схему строения хромосом. Основным направлением работ Ю. А. Филипченко была частная генетика животных и растений. Ему принадлежит разработка приемов скрещивания для улучшения пород домашнего скота. Он первым начал читать курс лекций по генетике в университетах России, создал первую кафедру генетики в Петроградском университете и первую Лабораторию генетики при Академии наук. Н. И. Вавилов является гордостью отечественной генетики. Основные научные работы его посвящены вопросам генетики, селекции и формообразования у растений. Н. И. Вавилов был организатором и руководителем многочисленных экспедиций по изучению мировых растительных ресурсов (Иран, Афганистан, страны Средиземноморья, Эфиопия, Япония, Корея, Северная, Центральная и Южная Америка и др.). Им установлены очаги формообразования, или центры происхождения культурных растений, обоснованы принципы селекции растений, сформулирован закон гомологических рядов в наследственной изменчивости, создана мировая коллекция культурных растений, создано учение об иммунитете растений, разработаны принципы селекции растений. Характерной чертой исследований Н. И. Вавилова была широта и глубина теоретических обобщений. Его огромное литературное наследие является достоянием мировой науки. Существенную роль в развитии отечественной генетики сыграл С. С. Четвериков (1880–1959). Основное направление его работ – генетика популяций. Им сформулированы основные положения популяционной генетики, принцип насыщенности природных популяций большим количеством мутаций, выведен закон равновесия при свободном скрещивании, установлена роль изоляции в дифференциации вида, а также значения естественного отбора в процессе эволюции. Благодаря заслугам С. С. Четверикова, а также последующим работам Н. П. Дубинина, Д. Д. Ромашова, Ю. М. Оленова и С. М. Гершензона в нашей стране, Р. Фишера и Д. Холдейна в Англии, С. Райта и Ф.Г. Добжанского в США, а также Н. В. Тимофеева-Ресовского (жившего в те годы в Германии), генетика оказалась прочно связанной с эволюционным учением и стала, по существу, фундаментом синтетической теории эволюции. Третий этап развития генетики (1925–1944) ознаменован в первую очередь открытием индуцированного мутагенеза. В 1925 г. советский микробиолог и генетик Г. А. Надсон совместно со своим учеником Г. С. Филипповым продемонстрировали появление мутантов у низших грибов под влиянием рентгеновских лучей. Однако в работах Г. А. Надсона мутационный процесс не был проанализирован количественными методами, а также в силу слабой изученности используемых объектов, его исследования не привлекли должного внимания. Только через два года глубоко продуманный и методически безупречный эксперимент Г. Меллера (1927 г.) на дрозофиле и Л. Стадлера (1928 г.) на ячмене и кукурузе полностью доказали возможность получения мутаций под действием радиации. Вслед за этими работами была продемонстрирована мутагенная активность ионизирующего излучения (рентгеновских лучей, альфа-, бета- и гамма-излучения и др.) в отношении различных объектов, в результате чего экспериментальный мутагенез стал мощным инструментом в генетических исследованиях. Несколько позже была обнаружена мутагенная активность ряда химических веществ. Первые опыты по химическому мутагенезу были проведены в 1930-х гг. отечественными учеными В.В. Сахаровым, М. Е. Лобашевым и С. М. Гершензоном, а затем в середине 1940-х гг. И. А. Рапопортом и английским генетиком Ш. Ауэрбах. В 1937 г. А. Блексли и О. Эвери было открыто действие колхицина, а в 1941 г. А. Холлендер с сотрудниками продемонстрировали мутагенное действие ультрафиолетового света. Исследования в области экспериментального мутагенеза привели к быстрому прогрессу в познании механизмов мутационного процесса и к выяснению ряда вопросов, касающихся тонкого строения гена. Из наиболее интересных работ в этом направлении можно назвать исследования А. С. Серебровского (1892–1948) и его учеников Н. П. Дубинина, И. И. Агола, А. Е. Гайсиновича и др., касающиеся доказательства сложного строения и дробимости гена и послужившие началом изучения его внутреннего строения. Тогда же начали интенсивно развиваться частные разделы генетики. Широко развернулись работы по генетике и генетическим основам селекции растений, проводимые П. М. Жуковским, Г. Д. Карпеченко, А. Р. Жебраком, Н. В. Цициным, А. А. Сапегиным и Л. Д. Делоне. В области генетики животных больших успехов добились А. С. Серебровский, М. Ф. Иванов, Б. Л. Астауров, Я. Л. Гелембоцкий, Д. К. Беляев, М. М. Завадский, Б. Л. Астауров. Ученые С. Г. Левит и С. Н. Давиденков организовали проведение широких исследований по генетике человека, а С. Н. Ардашников, А. А. Прокофьева-Бель-говская и В. П. Эфроимсон по медицинской генетике. Во второй половине 30-х гг. была заложена основа бурного развития биохимической генетики. Эксперименты Дж. Бидла и Б. Эфрусси (1935 г.) показали, что рецессивные мутации у дрозофилы могут приводить к блокированию разных участков в пути синтеза глазного пигмента. Еще больших успехов добились Дж. Бидл и Э. Тейтем (1941 г.), используя в качестве объекта гриб Neurospora crassа. Эти ученые установили, что каждый ген в организме определяет синтез одного конкретного фермента, и сформулировали концепцию «один ген – один фермент». Несмотря на революционный характер этой работы, ее еще нельзя отнести к эпохе современной генетики, так как, оперируя понятием «ген», Дж. Бидл и Э. Тейтем еще не имели представления об его материальной основе, «ген» для них оставался умозрительным понятием, что было характерно для периода «классической генетики», продолжавшегося до 1944 г., когда впервые была установлена природа наследственного материала. В начале 40-х гг. начала активно развиваться также генетика бактерий и бактериофагов. Был установлен факт возникновения спонтанных мутаций у бактерий (С. Луриа и М. Дельбрюк, 1943), а также доказано существование у них процесса рекомбинации (Дж. Ледерберг и Э. Тейтем, 1946 г.). В это же время были обнаружены мутации у бактериофагов (А. Херши и М. Дельбрюк, 1946 г.). Особенно благоприятным объектом для изучения генетических процессов оказались бактерии Escherichia coli, а также их бактериофаги (λ, Т2 и Т4, φХ 174 и др.), высокая скорость размножения которых и простота генома позволили генетикам выйти на новый уровень исследований. Четвертый этап развития генетики (1944–1960). Наиболее характерными чертами этого периода развития генетики является переход на новый молекулярный уровень изучения структуры генетического материала и закономерностей передачи наследственных признаков. Решительный поворот в понимании роли нуклеиновых кислот как носителей наследственной информации произошел в 1944 г., когда бактериолог О. Эвери, Ч. Мак Леод и М. Мак Карти проверили работу Ф. Гриффита, описавшего процесс трансформации у пневмококков, и затем продемонстрировали, что носителем наследственной информации является ДНК. В 1952 г. А. Херши и М. Чейз представили окончательное подтверждение роли генетической роли ДНК на примере бактериофага Т2 Доказательство участия РНК в передаче наследственных признаков было представлено в 1956–1957 гг. Г. Френкель-Конратом и Р. Вильямсом. Установление А. Астбери, М. Уилкинсом и Р. Франклин (1950–1952) пространственной организации молекулы ДНК и особенностей ее химического строения (Э. Чаргафф, 1950 г. и позже) создали основу для окончательного выяснения ее структуры. В 1953 г. Дж. Уотсон и Ф. Крик показали, что молекула ДНК имеет форму двойной спирали, а также сформулировали принцип комплементарности отдельных цепей в молекуле. Именно комплементарность была положена в основу объяснения механизма репликации ДНК в последующем. В 1958 г. Ф. Крик сформулировал «центральную догму молекулярной биологии», согласно которой передача наследственной информации идет только в одном направлении, а именно от ДНК к РНК и от РНК к белку. Окончание четвертого этапа можно охарактеризовать, как накопление информации о тонком строении структур клетки, которые задействованы в генетических процессах. Так, в 1955 г. Г. Паладе были детально описаны рибосомы. В 1957 г. М. Хоглэнд, П. Замечник и М. Стефенсон обнаружили тРНК. В этом же году А. Корнберг описал новый фермент ДНК-полимеразу I. В 1960 г. Ф. Жакоб, С. Бреннер и М. Мезельсон обнаружили мРНК. Особенностью открытий того времени было то, что практически все они были сделаны с использованием в качестве объектов бактерий и бактериофагов. Эти работы стали мощным толчком дальнейшего развития молекулярной генетики и всей молекулярной биологии. Пятый этап развития генетики (1960–1990) можно охарактеризовать как период развития молекулярной генетики на основе накопленных ранее данных и появления новых основополагающих открытий. В 1960 г. одновременно тремя коллективами, возглавляемыми Дж. Гурвитцем, Н. Стивенсом и С. Вейссом, описан процесс транскрипции. В 1961 г. разработана модель оперона Ф. Жакобом и Ж. Моно, а в 1965– 1965 гг. М. Ниренбергом, Дж. Маттеи, С. Очоа и Г. Корана расшифрован генетический код. В это же время установлены молекулярные механизмы основных генетических процессов – репликации, транскрипции и трансляции. Второй особенностью этого периода является постепенное вовлечение в круг исследований новых, более сложных объектов – растений, животных и человека. Третьей особенностью является бурное развитие и применение в генетике методологий молекулярной биологии. Так, в 1970 г. Г. Смитом, Д. Натансом и В. Арбером были открыты рестриктазы, а Д. Балтимором в этом же году – обратная транскриптаза, что открыло путь к разработке технологии рекомбинантных ДНК. Годом рождения генетической инженерии считается 1972 г., когда П. Берг впервые получил рекомбинантную ДНК in vitrо. В этом же году Г. Боером и С. Кохеном осуществлено клонирование первого гена в составе плазмидного вектора. Открытия этого периода можно назвать «революцией рекомбинантных ДНК». В 1977 г. А. Максам, В. Гилберт и Ф. Сэнгер разработали методику секвенирования ДНК. В 1986 г. К. Мюллис – методологию полимеразной цепной реакции и др. Шестой этап развития генетики (1990– настоящее время.). Создание новых высоко технологичных методологий исследования генетического аппарата различных организмов послужило базой для начала новой эры развития генетики, характерной чертой которой является решение глобальных генетических задач, в том числе связанных со здоровьем человека. Этот этап развития генетики знаменуется началом исследований по расшифровке генома человека (проект «Геном человека», 1990 г.), разработкой новых подходов идентификации генов, лечения наследственных заболеваний человека, в том числе с помощью генотерапии, совершенствованием методологии получения трансгенных животных и растений и др. Вполне заслужено 1990 г. может быть отнесен к эпохе «геномной» революции, очевидцами которой мы являемся. Понятие о наследственности и изменчивости.Понятие наследственности. Наследственная информация о признаках организма закодирована в генах, которые являются участками более сложных образований – хромосом. Ведущая роль в передаче наследственной информации принадлежит материальной структуре – ДНК (дезоксирибонуклеиновая кислота). ДНК – сложный биологический полимер. Он состоит из длинной цепочки нуклеотидов. В состав каждого нуклеотида входят: остаток фосфорной кислоты, пентозный сахар дезоксирибоза, одно из азотистых оснований (пуриновые – аденин, гуанин; пиримидиновые – тимин, цитозин). ДНК способна самовоспроизводиться и сохранять наследственную информацию, закодированную в ней в виде последовательности чередования нуклеотидных оснований, во множестве поколений клеток, образующихся в онтогенезе многоклеточного организма. Реализация наследственной информации, закодированной в молекулах ДНК, осуществляется в процессе синтеза белка Хромосомная теория наследственности. Согласно хромосомной теории наследственности Моргана, гены, расположенные в одной и той же хромосоме, составляют группу сцепления и наследуются совместно (сцепленно) в соответствии с поведением этой хромосомы при мейозе и при оплодотворении. Сцепленное наследование ограничивает третье правило Менделя (правило независимого наследования признаков). Число групп сцепления у всех видов организмов соответствует гаплоидному набору хромосом. Сцепление может быть полным и неполным. Сцепление генов может нарушаться в результате рекомбинации, происходящей путем кроссинговера в процессе мейоза. Гомологичные хромосомы при этом обмениваются идентичными участками (генами), что обеспечивает расхождение сцепленных генов в разные гаметы. Чем дальше друг от друга отстоят гены в хромосоме, тем меньше силы их сцепления, тем больше частота кроссинговера, чаще нарушается сцепление и наоборот. Кроссинговер приводит к новому сочетанию генов в хромосомах, это является одной из причин комбинативной изменчивости (появление потомков с новым сочетанием признаков родителей). Гаметы и особи с новым сочетанием, появившиеся в результате кроссинговера, называются кроссоверами или рекомбинантами, а само явление – генетической рекомбинацией. Наличие сцепления выясняется по результатам анализирующего скрещивания. Если сцепления нет (независимое наследование признаков), то при скрещивании дигетерозиготы с дирецессивной формой (анализирующее скрещивание) среди потомков с равной частотой встречаются особи с родительскими сочетаниями признаков и с новыми их сочетаниями, т. е. в соотношении 1 : 1 : 1 : 1. При неполном сцеплении среди потомков анализирующего дигибридного скрещивания больше половины их имеют родительские сочетания признаков и меньше половины – новые сочетания, т. е. кроссоверов всегда будет меньше 50 % В случае сцепления число кроссоверных особей служит для определения расстояния между данными генами в хромосомах. Частота кроссинговера определяется в процентах и отражает силу сцепления между генами, которая и показывает расстояние между генами. 100, n n Частота кроссинговера 1 где n1 – число кроссоверов; n – общее число потомков. За единицу расстояния (перекреста) между генами в хромосоме принят 1 % кроссинговера. Эта единица называется морганидой. На основании анализа частоты кроссинговера между генами к настоящему времени для многих видов животных и растений построены карты хромосом. Картой хромосом называется план расположения генов в хромосоме. Составление карт хромосом начинается с установления группы сцепления, т.е. определения локализации группы сцепления генов в конкретной паре гомологичных хромосом. Для этого проводят скрещивание гетерозигот из F1 c гомозиготным рецессивом и сопоставление соотношения в Fа; для сцепленного наследования при анализирующем скрещивании характерно преобладание в фенотипических классах Fа особей с сочетанием родительских признаков и меньшим числом рекомбинантов. Для картирования генов в хромосоме необходимо, как минимум, установление сцепления трех и более неаллельных генов. Понятие изменчивости. Изменчивость — способность живых организмов приобретать новые признаки и свойства. Благодаря изменчивости, организмы могут приспосабливаться к изменяющимся условиям среды обитания. Различают две основные формы изменчивости: наследственная и ненаследственная. Наследственная, или генотипическая, изменчивость — изменения признаков организма, обусловленные изменением генотипа. Она, в свою очередь, подразделяется на комбинативную и мутационную. Комбинативная изменчивость возникает вследствие перекомбинации наследственного материала (генов и хромосом) во время гаметогенеза и полового размножения. Мутационная изменчивость возникает в результате изменения структуры наследственного материала. Ненаследственная,или фенотипическая, или модификационная, изменчивость — изменения признаков организма, не обусловленные изменением генотипа. Мутации.Мутации — это стойкие внезапно возникшие изменения структуры наследственного материала на различных уровнях его организации, приводящие к изменению тех или иных признаков организма. Генные мутации возникают при изменении структуры молекулы ДНК в следующих случаях: – при замене пар нуклеотидов (одной или нескольких) на другие; – при выпадении пар нуклеотидов; – при включении (добавлении) пар нуклеотидов в цепочку ДНК. Мутации типа замены пар оснований, например, А-Т на Г-Ц, или наоборот, могут привести к изменению смысла кодона, и тогда в соответствующем месте полипептида произойдет замена одной аминокислоты на другую Мутации типа выпадения или вставки оснований, при которых происходит сдвиг рамки считывания, приводят, как правило, к более существенным последствиям, так как дистальнее точки мутации меняется вся последовательность аминокислот в полипептиде. В том случае, когда выпадет или вставится в ДНК триплет оснований (кодон) изменения в полипептиде будут не столь значительны, так как выпадет или вставится одна аминокислота Возвратное и анализирующее скрещивание.Скрещивание особей, различающихся по одной паре признаков, называется моногибридным. Проводя моногибридное скрещивание, Г.Мендель открыл два закона наследования качественных признаков. Первый закон Г.Менделя – при скрещивании двух гомозиготных особей, отличающихся друг от друга по состоянию развития одного признака, потомки первого поколения (F1) получаются единообразными как по фенотипу (с доминантным признаком), так и по генотипу (гетерозиготными). Первый закон Г.Менделя еще известен как правило доминирования и единообразия гибридов первого поколения. Второй закон Г.Менделя, или правило расщепления – при скрещивании двух гомозиготных особей, отличающихся друг от друга по состоянию развития одного признака, среди потомков второго поколения (F2) наблюдается расщепление как по фенотипу (три части потомков получаются с доминантным признаком, одна часть – с рецессивным), так и по генотипу (1АА, 2Аа, 1аа) У сельскохозяйственных животных встречаются разные формы взаимодействия аллельных генов (разные формы доминирования признаков): полное доминирование, неполное доминирование или промежуточное наследование; кодоминирование, сверхдоминирование. Возвратными называют скрещивания, при которых гибрид повторно скрещивают с одной из родительских форм. Их применяют в двух случаях. 1. Для преодоления бесплодия гибридов первого поколения при отдаленной гибридизации; такое скрещивание можно представить в виде формулы; (АxБ)xБ. 2. Для усиления в гибридном потомстве желаемых свойств одной из родительских форм. Формула такого скрещивания: АxБ-> АБxБ -> АББxБ -> АБББxБ -> АББББxБ и т. д. Анализирующее скрещивание, аналитическое скрещивание, тест-кросс (test crossing, test cross) [греч. analysis — разложение, расчленение] — скрещивание гибридной особи с особью-«анализатором», гомозиготной по рецессивным аллелям. Потомки от А.с. обязательно несут один рецессивный аллель от «анализатора», на фоне которого должны проявиться аллели, полученные от анализируемого организма. Для А.с. (исключая случаи взаимодействия генов) характерно совпадение расщепления по фенотипу с расщеплением по генотипу среди потомков. Таким образом, А.с. позволяет определить генотип и соотношение гамет разного типа, образуемых анализируемой особью. Возвратное скрещивание, беккросс (backcross) — скрещивание гетерозиготных гибридов F1 (Аа) с гомозиготными особями, сходными по генотипу с родительскими формами (АА или аа). Метод возвратного скрещивания состоит в получении потомства в ряду поколений от скрещивания гетерозиготы (детей гомозиготных родителей, генетически отличающихся друг от друга) с одним из исходных гомозиготных родителей. Решение задач.У крупного рогатого скота сплошная окраска (ген «С») доминирует над пестрой (ген «с»). В популяции беспородного скота, насчитывающей 940 голов, 705 животных имели черно-пеструю масть и 235 – сплошную черную. Пользуясь формулой Харди-Вайнберга, определите частоту фенотипов и концентрацию генов «С» и «с». Если известны частоты разных фенотипов в популяции, то на основе формулы Харди-Вайнберга можно составить представление о распределении в ней соответствующих генотипов. Уравнение Харди-Вайнберга: P2 + 2pq +q2 = 1, где p – частота доминантной аллели; q – частота рецессивной аллели. 1. Условные обозначения генов Известно, что перстая окраска – рецессивный признак (ген «с»), поэтому генотип перстая «сс», а генотип нормальных особей «СС» и «Сс». 2.Определим частоту перстых «сс» и выразим ее в долях единицы: q2сс =705/940=0.75 3.Частота перстой окраски «с» составит: qc = 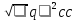 =0,86 =0,864. Определим частоту доминантной «С» на основе формулы pС + qс = 1 pС = 1 – qс = 1 – 0,86= 0,14 5. Определяем частоту гомозигот и гетерозигот среди фенотипически нор-мальных особей: р2СС = 0,0196 или 1,96 % 2рqСС = 2 * 0,86 * 0,14=0,2408 или 24,08 %. Ответ: генетическая структура популяции такова – СС – 1,96% Сс – 24,08 % сс – 75 % Выпишите и дайте объяснение всем терминам, встречавшимся вам при выполнении работы.ДНК это биологический полимер, состоящий из двух полинуклеотидных цепей, соединенных друг с другом. ДНК – полимер с очень большой молекулярной массой. Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие азотистые основания: аденин (А) или тимин (Т), цитозин (Ц) или гуанин (Г), пятиатомный сахар – пентозу – дезоксирибозу, по имени которой получила название и сама ДНК, а также остаток фосфорной кислоты. Хромосомы это органоиды клеточного ядра, являющиеся носителями генов и определяющие наследственные свойства клеток и организмов. Совокупность полного набора хромосом (число, размеры, форма и т.д.), присущего клеткам данного биологического вида, называют кариотипом. Мейоз (редукционное деление клетки) — деление, в процессе которого из одной диплоидной (2n) клетки получаются 4 гаплоидные (n) клетки. Так как у дочерних клеток происходит уменьшение (редукция) числа хромосом с 2n до n, такое деление названо редукционным. Кроссинговер - это явление в результате которого происходить обмен участками ДНК (генами) у гомологичных хромосом. Обмен осуществляется во время временного сближения гомологичных хромосом, что характерно в период формирования гамет или спор. Чем ближе изначально (до сближения) эти гены расположены друг к другу, тем меньше вероятность кроссинговера. Гомологи́чные хромосо́мы — пара хромосом с одинаковым набором генов и сходной морфологии в кариотипе диплоидного организма. Гаметы, или половые клетки, — репродуктивные клетки, имеющие гаплоидный (одинарный) набор хромосом и участвующие, в частности, в половом размножении. Генетика – наука о наследственности и изменчивости организмов. Наследственность – способность живых организмов передавать свои признаки и свойства, а также особенности развития из поколения в поколение. Изменчивость – способность живых организмов приобретать новые признаки и свойства. Признак – особенность строения на любом уровне организации или внешнее проявление действия гена. Свойство – любая функциональная особенность, в основе которой лежит один или несколько признаков. Альтернативные признаки – взаимоисключающие, контрастные признаки. Ген – участок молекулы ДНК, отвечающий за один признак, т.е. за структуру определенной молекулы белка. Локус – местоположение гена в хромосоме. Аллельные гены – гены, расположенные в одних и тех же локусах гомологичных хромосом и ответственные за развитие одного признака. Геном – совокупность генов, характерных для гаплоидного набора хромосом данного вида организмов. Генотип – совокупность взаимодействующих генов данного организма. Фенотип – совокупность всех признаков и свойств организма. Доминантный признак – преобладающий признак, проявляющийся в потомстве у гетерозиготных особей. Рецессивный признак – подавляемый, внешне исчезающий признак. Гомозигота – зигота, имеющая одинаковые аллели данного гена. В потомстве не дает расщепление. Полигибридное скрещивание – скрещивание форм, отличающихся друг от друга по трем и более парам альтернативных признаков. Анализирующее скрещивание – скрещивание гибридной особи с особью, гомозиготной по рецессивным аллелям. Неполное доминирование – случаи, когда гетерозиготные потомки имеют промежуточный фенотип. Кариотип – совокупность признаков хромосомного набора соматической клетки. Аутосомы – хромосомы, одинаковые у обоих полов. Половые хромосомы – хромосомы, по которым мужской и женский пол отличаются друг от друга. Гомогаметный – пол, который образуют гаметы, одинаковые по половой хромосоме (xx). Гетерогаметный – пол, который формирует гаметы, неодинаковые по половой хромосоме (xy). Список литературы.1. Бакай, А.В. Генетика / А.В. Бакай, И.И. Кочиш, Г.Г. Скрипниченко. – М.: Колос, 2006. – 448 с. 2. Инге-Вечтомов, С.Г. Генетика с основами селекции / С.Г. ИнгеВечтомов. – М.: Высшая школа, 1989 3. Казанцева, Н.П. Сборник задач по генетике. Учебно-методическое пособие для студентов зооинженерного и ветеринарного факультетов / Н.П. Казанцева. – Ижевск, 1998 4. Казанцева, Н.П. Словарь терминов по генетике, основам селекции и биотехнологии / Н.П. Казанцева. – Ижевск, 2006 5. Казанцева, Н.П. Практикум по генетике с основами биометрии / Н.П. Казанцева, А.И. Любимов, Е.Н. Мартынова, О.Г. Пушкарев. – Ижевск, 2005 6. Кайданов, Л.З. Генетика популяций / Л.З. Кайданов. – М.: Высшая школа, 1996 7. Карманова, Е.П. Практикум по генетике / Е.П. Карманова, А.Е. Болгов. – Петрозаводск, 2004 8. Крюков, А.М., Задачник по генетике животных / А.М. Крюков, Н.В Горбунова. – Пенза, 2005 |