химия. 15 лекция химия. Рнк), и дезоксирибонуклеиновые, содержащие дезоксирибозу (днк
 Скачать 1.01 Mb. Скачать 1.01 Mb.
|

7) нуклеотиды находятся в ионизированном состоянии. |
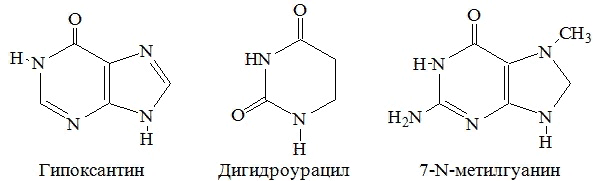
| Азотистое основание | Пентоза | Нуклеозид |
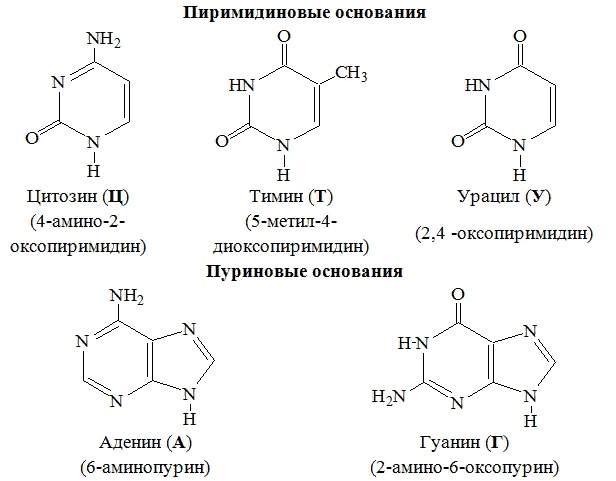
| Гуанин | Рибоза | Гуанозин |
| Аденин | Рибоза | Аденозин |
| Цитозин | Рибоза | Цитидин |
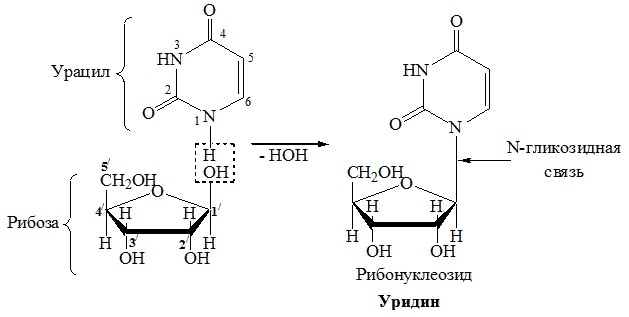
| Урацил | Рибоза | Уридин |
| Гуанин | Дезоксирибоза | Дезоксигуанозин |
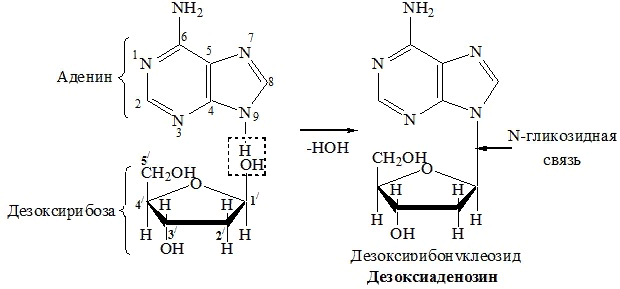
| Аденин | Дезоксирибоза | Дезоксиаденозин |
| Цитозин | Дезоксирибоза | Дезоксицитидин |
| Тимин | Дезоксирибоза | Тимидин |
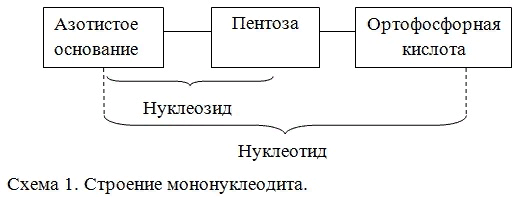
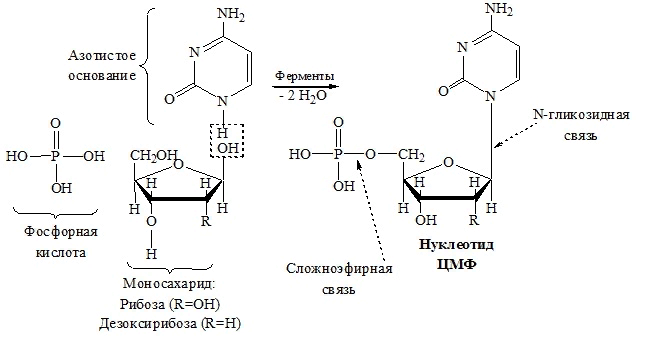 ассмотрим строение и составные части нуклеотида на примере образования цитидинмонофосфата (схема 4.)
ассмотрим строение и составные части нуклеотида на примере образования цитидинмонофосфата (схема 4.)В номенклатуре нуклеотидов используют два вида названий: одновключаетнаименованиенуклеозида с указаним положення в нем фосфатного остатка (например,цитидин-5'-монофосфат), другоестроится с добавлениемсуффикса-овая кислота к названиюостаткапиримидинового (например, 5'-цитидиловая кислота) или пуринового (например, 5'-гуаниловая кислота) оснований. Положение остатка фосфорной кислоты определяется местом разрыва фосфодиэфирной связи между соседними нуклеотидами.
Примеры некоторых названий нуклеотидов представлены в таблице 2.
Таблица 2.
Номенклатура нуклеотидов
| Названия нуклеотидов как монофосфатов | Названия нуклеотидов как кислот |
| Аденозин-5'-монофосфат (АМФ) | 5'-адениловая кислота |
| Гуанозин-5'-монофосфат (ГМФ) | 5'-гуаниловая кислота |
| Уридин-5'-монофосфат (УМФ) | 5'-уридиловая кислота |
| Цитидин-5'-монофосфат (ЦМФ) | 5'-цитидиловая кислота |
| Дезоксиаденозин-5'-монофосфат (дАМФ) | Дезоксиадениловая кислота |
| Дезоксигуанозин-5'-монофосфат (дГМФ) | Дезоксигуаниловая кислота |
| Дезоксицитидин-5'-монофосфат (дЦМФ) | Дезоксицитидиловая кислота |
| Тимидин-5'-монофосфат (ТМФ) | Тимидиловая кислота |
В биохимической литературе используют названия нуклеотидов как фосфатов с отражением этого признака в сокращенном коде (например, аденозин-5'-монофосфат - АМФ, аденозин-5'-дифосфат - АДФ, аденозин-5'-трифосфат - АТФ). Эти нуклеотиды способны взаимно превращаться путем присоединения одного или двух остатков фосфорной кислоты (фосфорилирование) или отщепления одного или двух остатков фосфорной кислоты (дефосфорилирование) (схема 5).
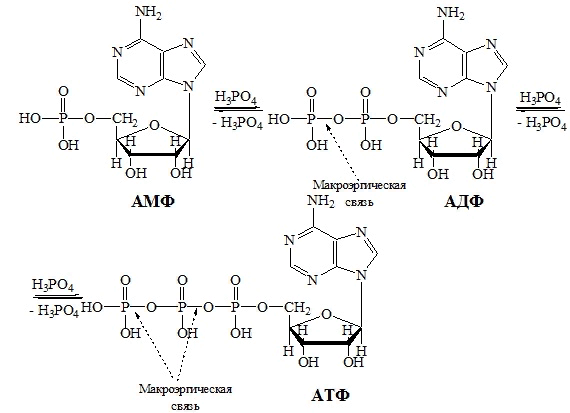
Схема 5. Фосфорилирование нуклеотидов.
При дефосфорилировании выделяется большое количество энергии, необходимое организму для протекания тех или иных биологических процессов. Связь О − Р между остатками ортофосфорной кислоты в молекулах нуклеозидполифосфатов носит название макроэргической связи. При ее гидролизе выделяется значительное количество энергии. Поэтому АТФ во многих биохимических процессах выполняет роль аккумулятора энергии.
Мононуклеотиды, динуклеотиды и их производные присутствуют в клетках также в свободном виде и играют важную роль в обмене веществ. В частности нуклеотидную структуру имеют многие коферменты, включая коферменты оксидоредуктаз, никотинамиднуклеотидные коферменты (НАД+, НАДФ+).
2.3. Принцип построения цепей ДНК и РНК
Нуклеиновые кислоты - высокомолекулярные соединения, в которых отдельные мононуклеотидные остатки связаны между собой в полинуклеотидных цепях 3' - 5' фосфодиэфирными связями. Стандартная запись нуклеотидной последовательности осуществляется в направлении от свободного 5' конца к свободному 3' концу. На схеме 6. приведено образование произвольного участка цепи РНК, включающего три мононуклеотидных остатка:
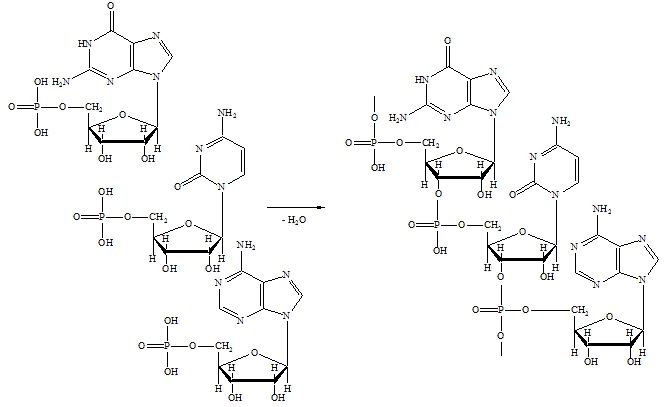
Схема 6. Образование фрагмента цепи РНК.
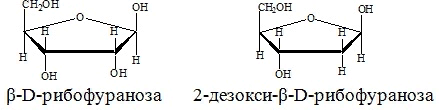
РНК, как правило, построены из одной полинуклеотидной цепи. Принцип построения цепи ДНК такой же, как и у РНК, с двумя исключениями: пентозным остатком в ДНК является 2-дезокси-Д-рибоза и в наборе азотистых оснований используется не тимин, а урацил.Схема 6. Образование фрагмента цепи РНК.
| Каждая нить ДНК представляет собой полинуклеотид. Это длинная цепь, в которой в строго определенном порядке расположены нуклеотиды. Молекула ДНК представляет собой две спирально закрученные одна вокруг другой нити.Вдоль молекулы ДНК могут быть уложены одна за другой тысячи белковых молекул. Молекулярная масса ДНК соответственно исключительно велика - она достигает десятков и даже сотен миллионов. |
3. Уровни структурной организации РНК и ДНК: первичная, вторичная и третичная структуры. Комплементарность азотистых оснований. Виды клеточных РНК. Их месторасположение, состав, размер, функции. Виды мутаций.
Нуклеиновые кислоты, подобно белкам, имеют несколько уровней структурной организации.
Первичная структура представляет собой определенную последовательность нуклеотидных звеньев в полинуклеотидной цепи РНК и ДНК. Она стабилизируется при помощи 3'- углеродным атомом пентозы одного нуклеотида и 5'-углеродным атомом пентозы другого мононуклеотида (схема 7.)
-
Ф
Остаток ортофосфорной кислоты
П
Остаток пентозы (рибозы или дезоксирибозы)
А.О.
Остаток азотистого основания
Схема 7. Первичная структура нуклеиновых кислот.
Для установления первичной структуры удобными объектами исследования оказались молекулы РНК. Известны три типа РНК: транспортная РНК (тРНК), информационная или матричная РНК (иРНК) и рибосомальная РНК (рРНК).Информационные РНК выполняют функцию матрицы белкового синтеза в рибосомах. Рибосомальные РНК выполняют роль структурных компонентов рибосом. Транспортные РНК участвуют в транспортировке α-аминокислот из цитоплазмы в рибосомы и в переводе нуклеотидной последовательности в аминокислотную последовательность. К настоящему времени удалось определить первичную структуру почти всех тРНК, ряда молекул рРНК, ряда вирусных РНК, в состав которых входят сотни и тысячи нуклеотидных остатков.Достигнуты большие успехи и в установлении первичной структуры ДНК.
 | Вторичная структура нуклеиновых кислот представляет собой пространственную ориентацию полинуклеотидных цепей в молекуле. Подавляющее большинство РНК живых существ характеризуются одноцепочным строением (рис.2). Однако у некоторых вирусов встречаются двухцепочечные молекулы РНК, выполняющие роль носителя генетической информации, Кроме того, двухцепочечные РНК образуются в процессе размножения тех вирусов, которые содержат одноцепочечную РНК. |
| Рис.2.Вторичная структура РНК |
Каждая молекула тРНК обладает постоянной вторичной структурой. Она имеет форму двумерного клеверного листа и состоит из спиральных участков, образованных нуклеотидами одной и той же цепи, и
| | расположенных между ними одноцепочечных петель. Каждая петля имеет свою функцию: например, одна является местом взаимодействия тРНК с рибосомами, другая служит для связывания с аминокислотами, третья - с ферментам и т.д. На рисунке 2. показана структура тРНК. Менее охарактеризована вторичная структура матричных и рибосомных РНК. |
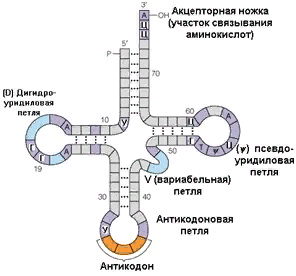 Рис.3. Вторичная структура транспортной РНК. |
Форма вторичной структуры молекулы ДНК - двойная спираль - изображена на рисунке 4. Впервые, в 1953 году, вторичную структуру ДНК в виде модели из двойной спирали описали английский биохимик Фрэнсис Крик и американский биохимик Джеймс Уотсон.
| | Согласно модели Уотсона - Крика молекула ДНК состоит из двух полинуклеотидных цепей, правозакрученных вокруг общей оси с образованием двойной спирали, имеющей диаметр 1,8 - 2,0 нм. Каждый виток спирали содержит 10 пар оснований и равен 3,4 нм. Две полинуклеотидные цепи антипараллельны друг другу, т.е. направления образования фосфодиэфирных связей в них противоположны: в одной цепи 5' → 3', в другой цепи 3' → 5'. |
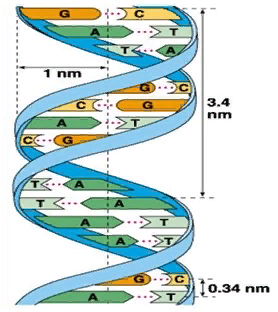 Рис. 4. Вторичная структура ДНК. |
Пиримидиновые и пуриновые основания направлены внутрь спирали. Формированию вторичной структуры способствуют водородные связи, которые образуются за счет специфического взаимодействия аденинового остатка одной цепи и тиминового остатка другой цепи (АТ) или гуанинового остатка одной цепи и цитозинового остатка другой цепи (ГЦ). Таким образом, последовательность нуклеотидов в одной цепи молекулы ДНК предопределяет нуклеотидную последовательность другой цепи.
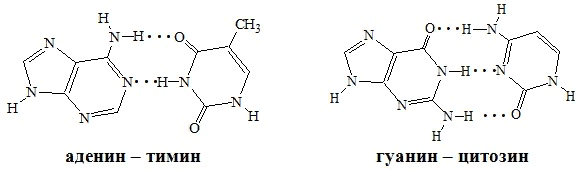
Основания, образующие пару, комплементарны друг другу, т.е. соответствуют друг другу по размерам и пространственной структуре, между
| | ними очень легко возникают водородные связи, которые являются основным условием стабильности биологической структуры. В комплементарной паре АТ имеется две водородные связи, а в комплементарной паре ГЦ - три такие связи (рисунок 5). Комплементарность цепей составляет химическую основу |
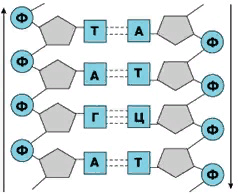 Рис.5.Участок двойной спирали ДНК |
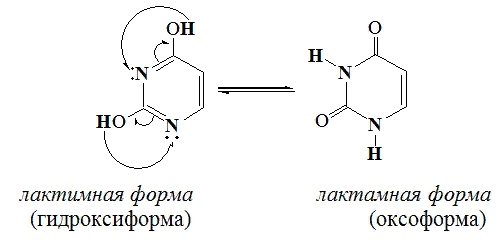
важнейшей функции ДНК - хранения и передачи наследственных признаков. Сохранность нуклеотидной первичной структуры является залогом безошибочной передачи генетической информации. Однако, под воздействием внешних факторов (мутагенов), нуклеотидная последовательность ДНК может быть нарушена и в организме возникают явления, называемые мутациями (от лат. mutatio - изменение), т.е. происходят стойкие изменения наследственных признаков К мутагенам относится, например, действие ионизирующего излучения, а также ряда химических веществ ( гидроксиламинNH2OH, азотистая кислота HNO2 и др.). Например, если на аденозин подействовать азотистой кислотой, то в результате реакции дезаминирования аминогруппа в адениновом основании превращается в гидроксильную группу. Результатом этой реакции будет превращение аденозина в другой нуклеозид - инозин, содержащий гипоксантин. Это может привести к замене в ДНК комплементарнойпары оснований, так как адениловый нуклеотид комплементарен тимину, а образовавшийся инозин может образовывать комплементарную пару только с цитозином. Замена одной пары азотистых оснований на другую - наиболее распространенный вид мутаций. Она передается при «переписывании» генетического кода с ДНК на РНК и приводит в итоге к изменению аминокислотной последовательности в синтезируемом белке. При накоплении мутаций возрастает число ошибок в биосинтезе белка.
Мутации под воздействием различных химических соединений имеют большое значение для управления наследственностью с целью ее улучшения. Результаты этих исследований нашли применение в селекциисельскохозяйственных культур, в создании штаммов микроорганизмов, производящих антибиотики, витамины.
Третичная структура нуклеиновых кислот.
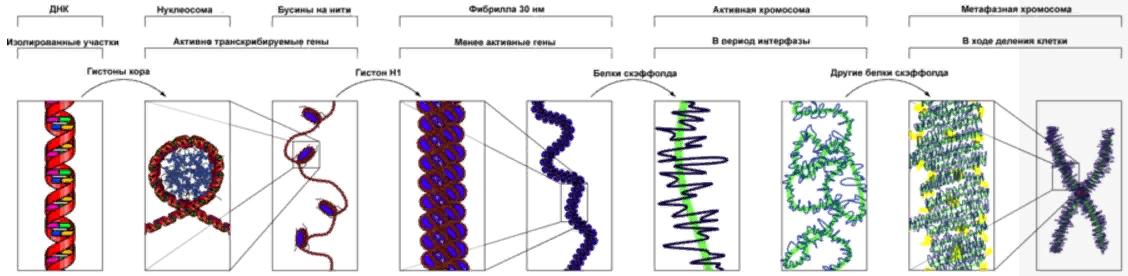
| | Двухцепочечная спираль ДНК в пространстве подвергается дальнейшей укладке в определенную третичную структуру -суперспираль(рисунок 6), или открытую кольцевую форму и занимает всего 1/5 объема клетки. Например, длина ДНК хромосомы человека достигает 8 см, а упакована так, что умещается в хромосоме с длиной 5 нм |
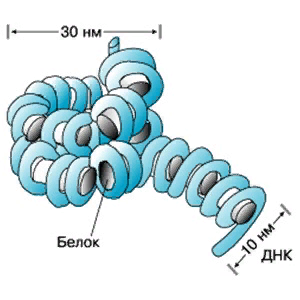 Рис. 6. Третичная структура ДНК (суперспираль). |
| | Суперспиральная конформация ДНК характерна для хромосом высших организмов (рисунок 7).Подобная третичная структура стабилизируется за счет ковалентных связей с остатками аминокислот, входящих в состав тех белков, которые образуют нуклеопротеидный комплекс. Существуют ферменты, которые способны увеличивать или уменьшать число витков, т.е. регулировать степень суперспиральности. Значение фактора суперспиральности ДНК чрезвычайно велико, так как некоторые белки и ферменты не связываются с ДНК, если она находится не в суперспиральнойконформации. |
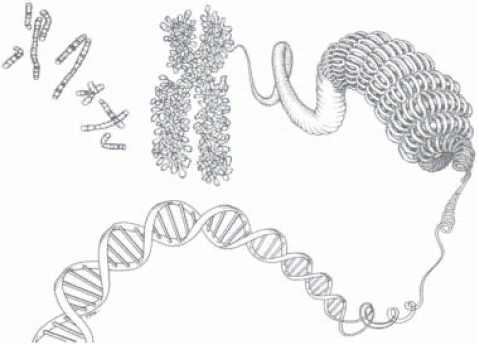 Рис.7. ДНК в составе хромосом |
В формировании третичной структуры РНК важную роль играют белки. При этом возникают большие внутриклеточные частицы - рибосомы, информосомы. Исключение составляют небольние по размерам транспортные РНК (70-80 мононуклеотидов). Третичная структура тРНК отличается от плоской структуры «клеверного листка» большой компактностью, образованной за счет складывания различных частей молекулы.
Биологическая роль РНК и ДНК.
Особенности химического строения нуклеиновых кислот обеспечивают возможность хранения, переноса в цитоплазму и передачи по наследству дочерним клеткам информации о структуре белковых молекул, которые синтезируются в каждой клетке.
Биологическая роль нуклеиновых кислот начала выясняться в конце 40-х ― начале 50-х годов, когда впервые было выяснено, что ДНК, взятая у одной разновидности бактерий и введенная в другую разновидность, заставляет последнюю производить потомство с признаками, имеющимися у первой разновидности. Отсюда вытекало, что вместе с ДНК была перенесена наследственная информация - каким-то образом закодированный приказ строить белковые молекулы определенного типа. Эти работы стали исходной точкой быстрого прогресса в области молекулярной генетики, приближающего нас к познанию процесса синтеза белка в клетках, размножения клеток путем деления и в конечном итоге воспроизведения всего сложного животного или растительного организма в том виде, который характерен для родителей этого организма. Подробное обсуждение этих проблем увело бы нас далеко в область биохимии, в общих же чертах роль ДНК и РНК выглядит следующим образом. Молекулы ДНК находятся в клеточных ядрах, они содержат наследственную информацию в виде различной последовательности нуклеотидов. ДНК играет роль матрицы, с которой отпечатываются копии молекул РНК, непосредственно участвующих в синтезе белков. Таким образом, молекулы РНК служат передатчиками от ДНК к местам клетки, где непосредственно осуществляется синтез белка.Белки обусловливают большинство свойств и признаков клеток. Понятно поэтому, что стабильность структуры нуклеиновых кислот - важнейшее условие нормальной жизнедеятельности клеток и организма в целом. Любые изменения строения нуклеиновых кислот влекут за собой изменения структуры клеток или активности физиологических процессов в них, влияя таким образом на жизнеспособность организмов.
| | Впервые модель молекулы ДНК была предложена в 1953 г. американским ученым Дж. Уотсоном и англичанином Ф. Криком на основе данных Э. Чаргаффа о соотношении пуриновых и пиримидиновых оснований молекул ДНК и результатов рентгеноструктурного анализа, полученных М. Уилкинсом и Р. Франклин. |
 Джеймс Уотсон и Фрэнсис Крик |
За разработку двухспиральной модели молекулы ДНК Джеймс Уотсон,ФрэнсисКрики Морис Уилкинс были удостоены в 1962 г. Нобелевской премии по физиологии и медицине.
В последовавшие десятилетия на основе открытий Д.Уотсона и Ф. Крика были совершены важные прорывы в области биологии и медицины. Открытие двойной спирали дало толчок последующему полувековому развитию молекулярной биологии, завершившемуся успешным осуществлением проекта «Геном человека».
В июне 2000 года Крейг Вентер и Фрэнсис Коллин (FrancisCollins), руководители проекта «Геном человека», осуществлявшегося в Национальных институтах здоровья США, объявили о событии, названном ими «первой сборкой генома человека». По существу, это была первая реконструкция полного генома человека, выполненная методом беспорядочной стрельбы. Несколькими месяцами позже, в феврале 2001 года, был опубликован первый предварительный набросок генома человека.
Можно считать, что на первом этапе проекта «Геном человека» была расшифрована книга жизни. На следующем этапе предстоит выяснить, что представляют собой все гены и как кодируемые ими белки объединяются, образуя биологический портрет человека. По оценкам ученых, на то, чтобы добыть все данные и понять все механизмы реализации генома человека, потребуется еще одно столетие.
Так или иначе, мы продвигаемся к пониманию полного генетического портрета человека, и это будет иметь грандиозные последствия для медицины и всего человечества.