Гистология (зрение). гиста зрение. Сенсорные системы. Органы чувств. Эмбриогенез и гистофизиология органа зрения
 Скачать 3.31 Mb. Скачать 3.31 Mb.
|

|
СЕНСОРНЫЕ СИСТЕМЫ. ОРГАНЫ ЧУВСТВ. ЭМБРИОГЕНЕЗ И ГИСТОФИЗИОЛОГИЯ ОРГАНА ЗРЕНИЯ Рецепторный аппарат глаза Рецепторный аппарат глаза представлен зрительной частью сетчатой оболочки (сетчатки). Внутренняя чувствительная оболочка глазного яблока, сетчатка (tunica interna sensoria bulbi, retina) состоит из наружного пигментного слоя (stratum pigmentosum) и внутреннего слоя нейросенсорных клеток (stratum nervosum) (рис. 12.5, а, б). Функционально выделяют заднюю большую зрительную часть сетчатки (pars optica retinae), меньшие части - ресничную, покрывающую ресничное тело (pars ciliares retinae), и радужковую, покрывающую заднюю поверхность радужки (pars iridica retina). В заднем полюсе глаза находится желтоватого цвета пятно (macula lutea) с небольшим углублением - центральной ямкой (fovea centralis). Свет входит в глаз через роговицу, водянистую влагу передней камеры, хрусталик, жидкость задней камеры, стекловидное тело и, пройдя через толщу всех слоев сетчатки, попадает на отростки нейросенсорных клеток, в наружных сегментах которых начинаются физиологические процессы возбуждения, фототрансдукции. Таким образом, сетчатка глаза человека относится к типу так называемых инвертированных органов, т. е. таких, в которых фоторецепторы направлены от света и образуют самые глубокие слои сетчатки, обращенные к слою пигментного эпителия. Фототрансдукция в наружном сегменте палочек или колбочек происходит при попадании света на клетку: 1. Далее происходит активация родопсина (смена цис-изоформы на транс-изоформу), которая по каскаду стимулирует белок трансдуцин (G-белок), который активирует фермент цГМФ фосфодиэстеразу.  2. Данный фермент катализирует превращение цГМФ в 5-ГМФ. 3. Снижение цГМФ ведет к закрытию натриевых каналов, что приводит к гиперполяризации фоторецептора. ! В темноте натриевые каналы открыты. ! При действии света они закрыты. 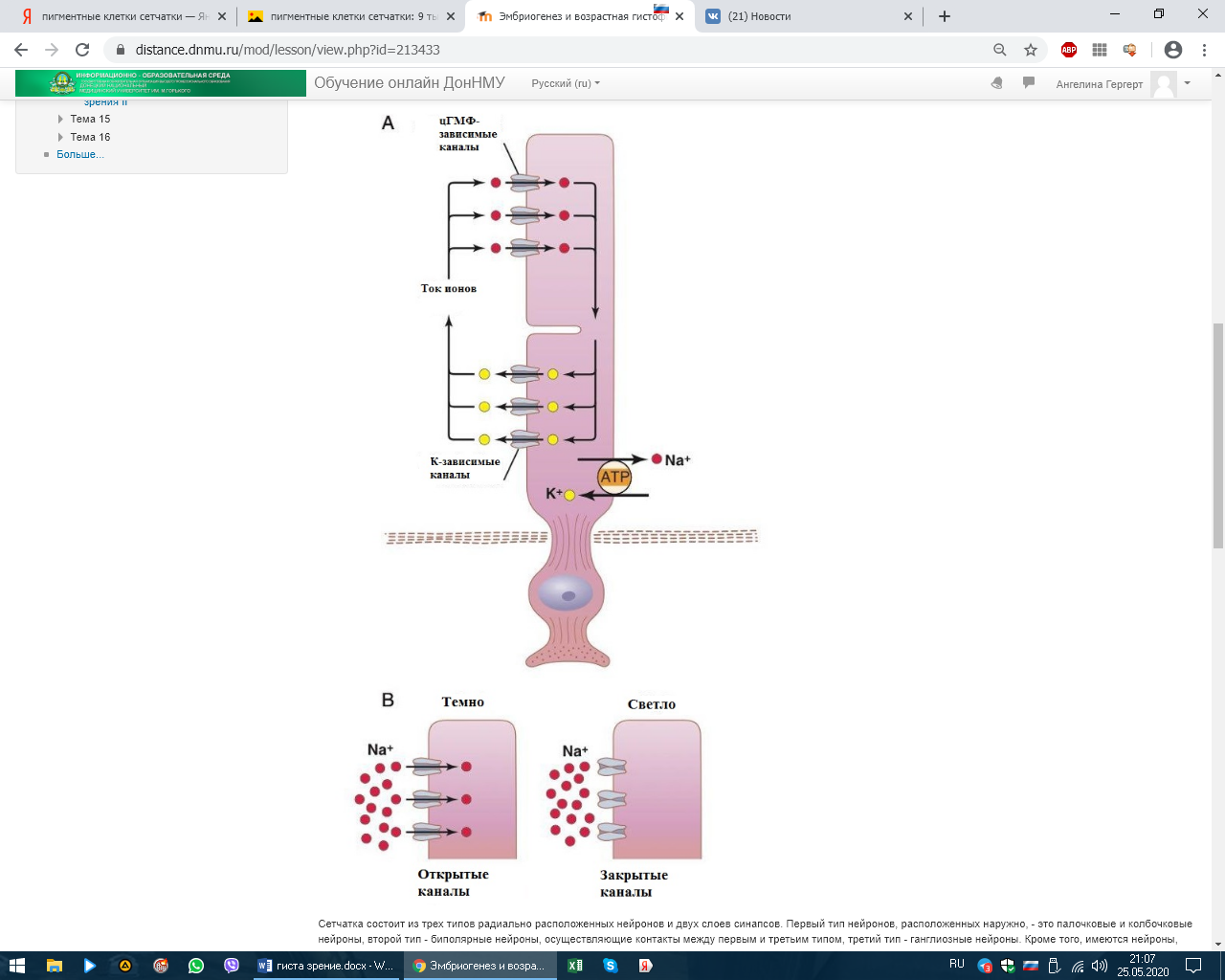 Сетчатка состоит из трех типов радиально расположенных нейронов и двух слоев синапсов. Первый тип нейронов, расположенных наружно, - это палочковые и колбочковые нейроны, второй тип - биполярные нейроны, осуществляющие контакты между первым и третьим типом, третий тип - ганглиозные нейроны. Кроме того, имеются нейроны, осуществляющие горизонтальные связи, - горизонтальные и амакринные. Наружный ядерный слой содержит тела палочковых и колбочковых нейронов, внутренний ядерный слой - тела биполярных, горизонтальных и амакринных нейронов, а слой ганглиозных клеток - тела ганглиозных и перемещенных амакринных нейронов (см. рис. 12.5). 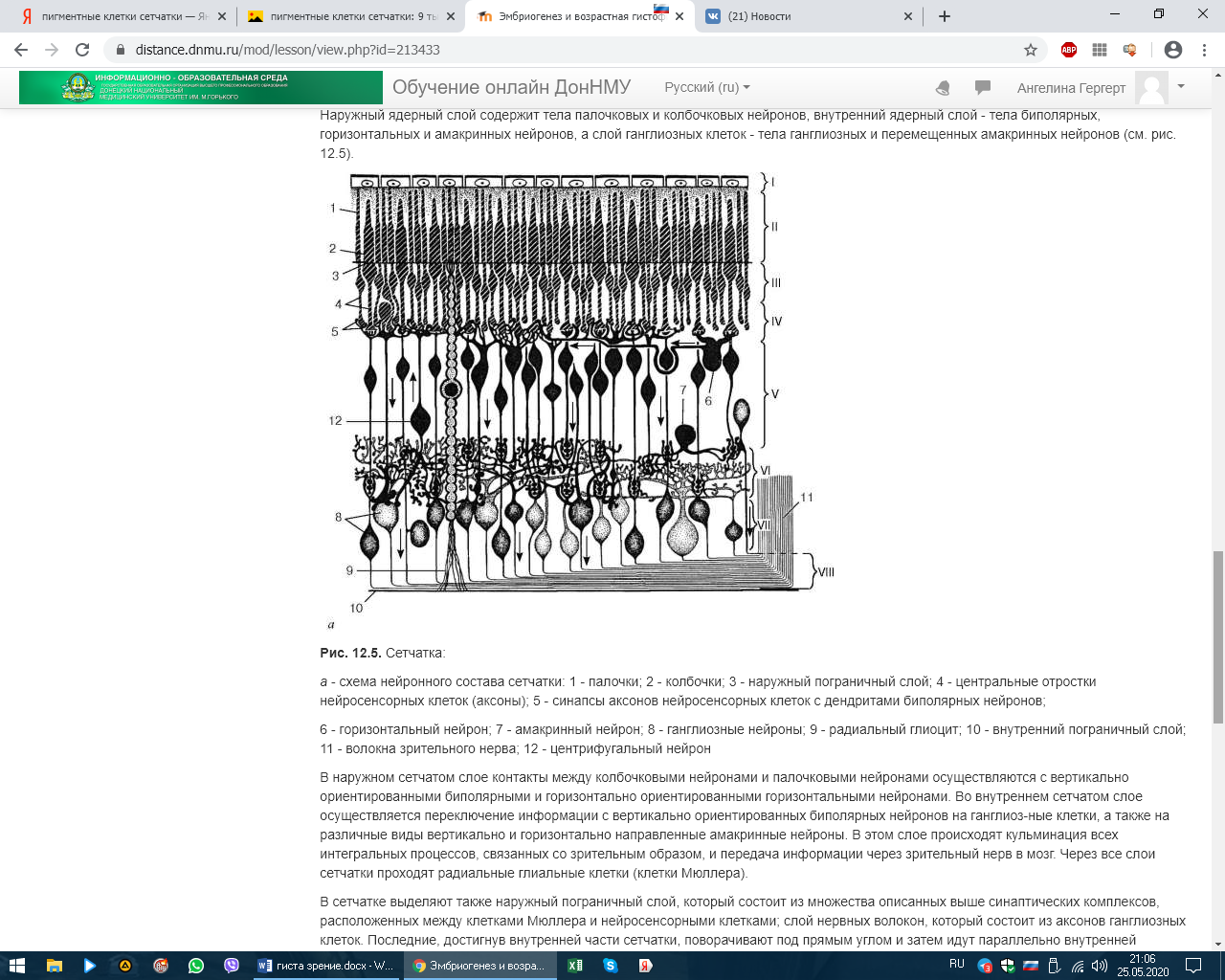 Рис. 12.5. Сетчатка: а - схема нейронного состава сетчатки: 1 - палочки; 2 - колбочки; 3 - наружный пограничный слой; 4 - центральные отростки нейросенсорных клеток (аксоны); 5 - синапсы аксонов нейросенсорных клеток с дендритами биполярных нейронов; 6 - горизонтальный нейрон; 7 - амакринный нейрон; 8 - ганглиозные нейроны; 9 - радиальный глиоцит; 10 - внутренний пограничный слой; 11 - волокна зрительного нерва; 12 - центрифугальный нейрон В наружном сетчатом слое контакты между колбочковыми нейронами и палочковыми нейронами осуществляются с вертикально ориентированными биполярными и горизонтально ориентированными горизонтальными нейронами. Во внутреннем сетчатом слое осуществляется переключение информации с вертикально ориентированных биполярных нейронов на ганглиоз-ные клетки, а также на различные виды вертикально и горизонтально направленные амакринные нейроны. В этом слое происходят кульминация всех интегральных процессов, связанных со зрительным образом, и передача информации через зрительный нерв в мозг. Через все слои сетчатки проходят радиальные глиальные клетки (клетки Мюллера). В сетчатке выделяют также наружный пограничный слой, который состоит из множества описанных выше синаптических комплексов, расположенных между клетками Мюллера и нейросенсорными клетками; слой нервных волокон, который состоит из аксонов ганглиозных клеток. Последние, достигнув внутренней части сетчатки, поворачивают под прямым углом и затем идут параллельно внутренней поверхности сетчатки к месту выхода зрительного нерва. Они не содержат миелина и не имеют шванновских оболочек, что обеспечивает их прозрачность. Внутренний пограничный слой представлен окончаниями отростков клеток Мюллера и их базальными мембранами.  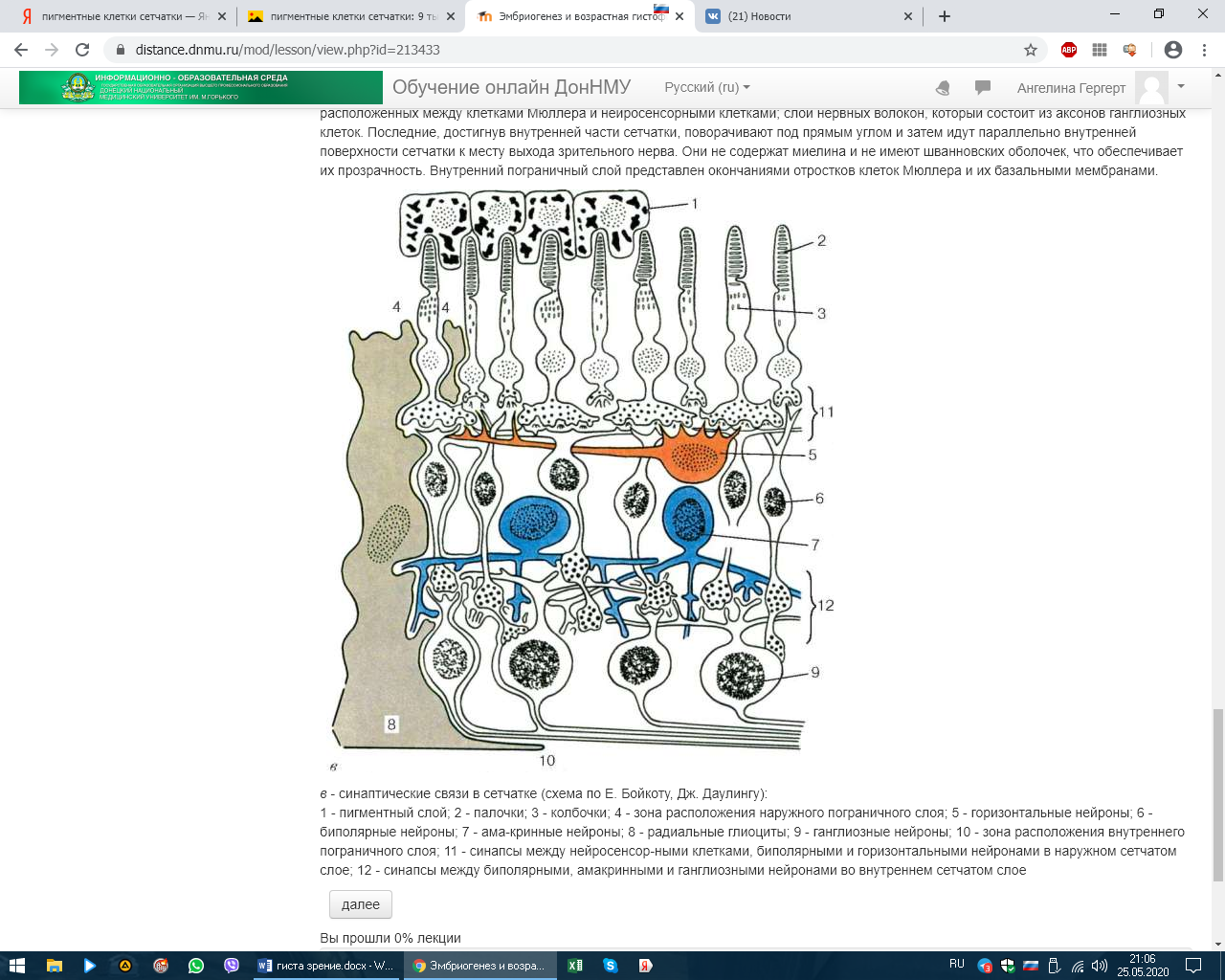 в - синаптические связи в сетчатке (схема по Е. Бойкоту, Дж. Даулингу): 1 - пигментный слой; 2 - палочки; 3 - колбочки; 4 - зона расположения наружного пограничного слоя; 5 - горизонтальные нейроны; 6 - биполярные нейроны; 7 - ама-кринные нейроны; 8 - радиальные глиоциты; 9 - ганглиозные нейроны; 10 - зона расположения внутреннего пограничного слоя; 11 - синапсы между нейросенсор-ными клетками, биполярными и горизонтальными нейронами в наружном сетчатом слое; 12 - синапсы между биполярными, амакринными и ганглиозными нейронами во внутреннем сетчатом слое При микроскопии задней стенки глаза латеральнее сосочка зрительного нерва выявили место наилучшего видения, где обнаружили только колбочковые фоторецепторные нейроны. В какой оболочке располагаются данные нейроны? – Сетчатой При микроскопии задней стенки глаза в одной из оболочек выявили клетки: палочковые, колбочковые, горизонтальные, амакринные, биполярные и ганглионарные. Укажите их клеточный тип. – Глиальные или Нервные Нейросенсорные клетки делятся на два типа: палочковые и колбочковые (см. рис. 12.5).  Рис. 12.5. Продолжение г, д - ультрамикроскопическое строение палочковой и колбочковой нейросенсорных клеток (схема по Ю. И. Афанасьеву): I - наружный сегмент; II - связующий отдел; III - внутренний сегмент; IV - перикарион; V - аксон. 1 - диски (в палочках) и полудиски (в колбочках); 2 - плазмолемма; 3 - базальные тельца ресничек; 4 - липидное тело; 5 - митохондрии; 6 - эндоплазматическая сеть; 7 - ядро; 8 - синапс Палочковые нейроны являются рецепторами сумеречного (ночного зрения), колбочковые нейроны - рецепторами дневного зрения. Морфологически нейросенсорные клетки представляют собой длинные цилиндрической формы клетки, которые имеют несколько отделов. Дистальная часть рецепторов - это видоизмененная ресничка. Наружный сегмент (палочка или колбочка) - содержит фоторецепторные мембраны, где и происходит поглощение света и начинается зрительное возбуждение. Наружный сегмент связан с внутренним сегментом соединительной ножкой - ресничкой (цилией). Во внутреннем сегменте находятся множество митохондрий и полирибосом, цистерны комплекса Гольджи и небольшое количество элементов гранулярной и гладкой эндоплазматической сети. В этом сегменте происходит синтез белка. Далее сужающаяся часть клетки заполнена микротрубочками (миоид), затем идет расширенная часть с ядром. Тело клетки, расположенное проксимальнее внутреннего сегмента, переходит в аксональный отросток, который формирует синапс с дендри-тами биполярных и горизонтальных нейронов. Однако палочковые клетки отличаются от колбочковых клеток (см. рис. 12.5, г, д). У палочковых нейронов наружный сегмент цилиндрической формы, а диаметр внутреннего сегмента равен диаметру наружного. Наружные сегменты колбочковых клеток обычно конические, а внутренний сегмент по диаметру значительно превосходит наружный. Наружный сегмент представляет собой стопку плоских мембранных мешочков - дисков, число которых доходит до 1000. В процессе эмбрионального развития диски палочек и колбочек образуются как складки - впячи-вания плазматической мембраны реснички. В палочках новообразование складок продолжается у основания наружного сегмента в течение всей жизни. Вновь появившиеся складки оттесняют старые в дистальном направлении. При этом диски отрываются от плазмо-леммы и превращаются в замкнутые структуры, полностью отделенные от плазмолеммы наружного сегмента. Отработанные диски фагоцитируются клетками пигментного эпителия. Дистальные диски колбочек так же, как у палочек, фагоцитируются пигментными клетками. Таким образом, фоторецепторный диск в наружном сегменте палочковых нейронов полностью отделен от плазматической мембраны. Он образован двумя фоторецепторными мембранами, соединенными по краям и внутри диска, на всем его протяжении имеется узкая щель. У края диска щель расширяется, образуется петля, внутренний диаметр которой составляет несколько десятков нанометров. Параметры диска: толщина - 15 нм, ширина внутридискового пространства - 1 нм, расстояние между дисками - междискового цитоплазматического пространства - 15 нм. У колбочек в наружном сегменте диски не замкнуты и внутридисковое пространство сообщается с внеклеточной средой (см. рис. 12.5, д). У них более крупное округлое и светлое ядро, чем у палочек. Во внутреннем сегменте колбочек имеется участок, называемый эллипсоидом, состоящий из липидной капли и скопления плотно прилегающих друг к другу митохондрий. От ядросодержащей части нейросенсорных клеток отходят центральные отростки - аксоны, которые образуют синапсы с дендритами биполярных и горизонтальных нейронов, а также с карликовыми и плоскими биполярными нейронами. Длина колбочек в центре желтого пятна около 75 мкм, толщина - 1-1,5 мкм. Толщина фоторецепторной мембраны наружного сегмента палочковых нейронов составляет около 7 нм. Основным белком фоторецепторной мембраны (до 95-98 % интегральных белков) является зрительный пигмент родопсин, который обеспечивает поглощение света и запускает фоторецеп-торный процесс. Зрительный пигмент представляет собой хромогликопротеид. Эта сложная молекула содержит одну хромофорную группу, две олигосахаридные цепочки и водонерастворимый мембранный белок опсин. Хромофорной группой зрительных пигментов служит ретиналь-1 (альдегид витамина А) или ретиналь-2 (альдегид витамина А2). Все зрительные пигменты, содержащие ретиналь-1, относятся к родопсинам, а содержащие ретиналь-2 - к порфиропсинам. Светочувствительная молекула зрительного пигмента при поглощении одного кванта света претерпевает ряд последовательных превращений, в результате которых обесцвечивается. Фотолиз родопсина запускает каскад реакций, в результате происходит гиперполяризация нейрона и уменьшение выделения медиатора. Среди колбочковых нейронов выделяют три типа, различающиеся зрительными пигментами с максимальной чувствительностью в длинноволновой (558 нм), средневолновой (531 нм) и коротковолновой (420 нм) части спектра. Один из пигментов - йодопсин - чувствителен к длинноволновой части спектра. Пигмент, чувствительный к коротковолновой части спектра, более сходен с родопсином. У человека гены, кодирующие пигмент коротковолновой части спектра и родопсина, находятся на длинном плече 3-й и 7-й хромосом и имеют сходство по структуре. Различные видимые нами цвета зависят от соотношения трех видов стимулируемых колбочковых нейронов. Отсутствие длинно- и средневолновых колбочковых нейронов обусловлено соответствующими изменениями гена на Х-хромосоме, которые определяют два типа дихромазии: протанопию и дейтеранопию. Протанопия - нарушение цветоощущения на красный цвет (ранее ошибочно называлось дальтонизмом). У Джона Дальтона благодаря последним достижениям молекулярной генетики выявлена дейтеранопия (нарушение цветоощущения на зеленый цвет). Горизонтальные нервные клетки (neuron horisontalis) располагаются в один или два ряда. Они отдают множество дендритов, которые контактируют с аксонами нейросенсорных клеток. Аксоны горизонтальных нейронов, имеющие горизонтальную ориентацию, могут тянуться на довольно значительном расстоянии и вступать в контакт с аксонами как палочковых, так и колбочковых нейронов. Передача возбуждения с горизонтальных клеток на синапсы нейросенсорной клетки и биполярного нейрона вызывает временную блокаду в передаче импульсов от фоторецепторов (эффект латерального торможения), что увеличивает контраст в зрительном восприятии. Биполярные нервные клетки (neuron bipolaris) соединяют палочковые и колбочковые нейроны с ганглиозными нейронами сетчатки. В центральной части сетчатки несколько палочковых нейронов соединяются с одним биполярным нейроном, а колбочковые нейроны контактируют в соотношении 1:1 или 1:2. Такое сочетание обеспечивает более высокую остроту цветового видения по сравнению с черно-белым. Биполярные нейроны имеют радиальную ориентацию. Различают несколько видов биполярных нейронов по строению, содержанию синаптических пузырьков и связям с фоторецепторами (например, биполярные нейроны палочки, биполярные нейроны колбочки). Биполярные клетки играют существенную роль в концентрации импульсов, получаемых от нейросенсорных клеток и затем передаваемых ганглиозным нейронам. Взаимоотношения биполярных нейронов с палочковыми и колбочко-выми нейронами различаются. Например, несколько палочковых клеток (15-20) в наружном сетчатом слое образуют синаптические связи с одним биполярным нейроном. Аксон последнего в составе внутреннего сетчатого слоя взаимодействует с различными типами амакринных нейронов, которые, в свою очередь, формируют синапсы с ганглиозным нейроном. Физиологический эффект заключается в ослаблении или усилении сигнала палочкового нейрона, что обусловливает чувствительность зрительной системы к единичному кванту света. Амакринные клетки относятся к интернейронам, которые осуществляют связь на втором синаптическом уровне вертикального пути: нейросенсорная клетка → биполярный нейрон → ганглиозный нейрон. Их синаптическая активность во внутреннем сетчатом слое проявляется в интеграции, модуляции, включении сигналов, идущих к ганглиозным нейронам. Эти клетки, как правило, не имеют аксонов, однако некоторые ама-кринные клетки содержат длинные аксоноподобные отростки. Синапсы амакринных клеток бывают химическими и электрическими. Например, дистальные дендриты амакринной клетки А образуют синапсы с аксонами палочковых биполярных нейронов, а проксимальные дендриты - с ган-глиозными нейронами. Более крупные дендриты А формируют электрические синапсы с аксонами колбочковых биполярных нейронов. В передаче нервного импульса от палочковых нейронов большую роль играют дофа-минергические и ГАМКергические амакринные клетки. Они ремодели-уют нервные импульсы и осуществляют обратную связь с палочковыми нейронами. Ганглиозные нейроны - наиболее крупные клетки сетчатки, имеющие большой диаметр аксонов, способных проводить электрические сигналы. В их цитоплазме хорошо выражено хроматофильное вещество. Они собирают информацию от всех слоев сетчатки как по вертикальным путям (нейросенсорные клетки → биполярные нейроны → ганглиозные нейроны), так и по латеральным путям (нейросенсорные клетки → горизонтальные нейроны → биполярные нейроны → амакринные нейроны → ганглиозные нейроны) и передают ее в мозг. Тела ганглиозных нейронов образуют ганглиозный слой (stratum ganglionicum), а их аксоны (более миллиона волокон) формируют внутренний слой нервных волокон (stratum neurofibrarum) и далее зрительный нерв. Ганглиозные нейроны гетероморфны. Они отличаются друг от друга по морфологическим и функциональным свойствам. Нейроглия. Три дифферона глиальных клеток найдены в сетчатке человека: клетки Мюллера (радиальные глиоциты), протоплазматические астроциты и микроглиоциты. Через все слои сетчатки проходят длинные, узкие радиальные глиальные клетки. Их удлиненное ядро лежит на уровне ядер биполярных нейронов. Базальные отростки клеток участвуют в образовании внутреннего, а апикальные отростки - наружного пограничного слоя. Клетки регулируют ионный состав окружающей нейроны среды, участвуют в процессах регенерации, играют опорную и трофическую роль. Пигментный слой, эпителий (stratum pigmentosum), наружный слой сетчатки - состоит из призматических полигональных пигментных клеток - пигментоцитов. Своими основаниями клетки располагаются на базальной мембране, которая входит в состав мембраны Бруха сосудистой оболочки. Общее количество пигментных клеток, содержащих коричневые гранулы меланина, варьирует от 4 до 6 млн. В центре желтого пятна пигментоциты более высокие, а на периферии они уплощаются, становятся шире. Апикальные части плазмолеммы пигментных клеток контактируют непосредственно с дистальной частью наружных сегментов нейросенсорных клеток. Апикальная поверхность пигментоцитов имеет два вида микроворсинок: длинные микроворсинки, которые располагаются между наружными сегментами нейросенсорных клеток, и короткие микроворсинки, которые взаимодействуют с концами наружных сегментов нейросенсорных клеток. Один пигментоцит контактирует с 30-45 наружными сегментами нейросен-сорных клеток, а вокруг одного наружного сегмента палочковых нейронов обнаруживается 3-7 отростков пигментоцитов, содержащих меланосомы, фагосомы и органеллы общего значения. В то же время вокруг наружного сегмента колбочкового нейрона - 30-40 отростков пигментоцитов, которые длиннее и не содержат органелл, за исключением меланосом. Фагосомы образуются в процессе фагоцитоза дисков наружных сегментов нейросен-сорных клеток. Наличие пигмента в отростках (меланосом) обусловливает поглощение 85-90 % света, попадающего в глаз. Под воздействием света меланосомы перемещаются в апикальные отростки пигментоцитов, а в темноте меланосомы возвращаются в перикарион. Это перемещение происходит с помощью микрофиламентов при участии гормона меланотропина. Пигментный эпителий, располагаясь вне сетчатки, выполняет ряд важных функций: оптическую защиту и экранирование от света; транспорт метаболитов, солей, кислорода и т. п. из сосудистой оболочки к нейросенсорным клеткам и обратно, фагоцитоз дисков наружных сегментов нейросенсорных клеток и доставку материала для постоянного обновления плазматической мембраны последних; участие в регуляции ионного состава в субретинальном пространстве. В пигментном эпителии велика опасность развития темновых и фотоокислительных деструктивных процессов. Все ферментативные и неферментативные звенья антиокислительной защиты присутствуют в клетках пигментного эпителия: пигментоциты участвуют в защитных реакциях, тормозящих перекисное окисление липидов с помощью ферментов микропероксисом и функциональных групп меланосом. Например, в них найдена высокая активность пероксидазы, как селе-нозависимой, так и селенонезависимой, и высокое содержание альфа-токоферола. Меланосомы в клетках пигментного эпителия, обладающие антиоксидантным свойством, служат специфическими участниками системы антиоксидантной защиты. Они эффективно связывают прооксидантные зоны (ионы железа) и не менее эффективно взаимодействуют с активными формами кислорода. 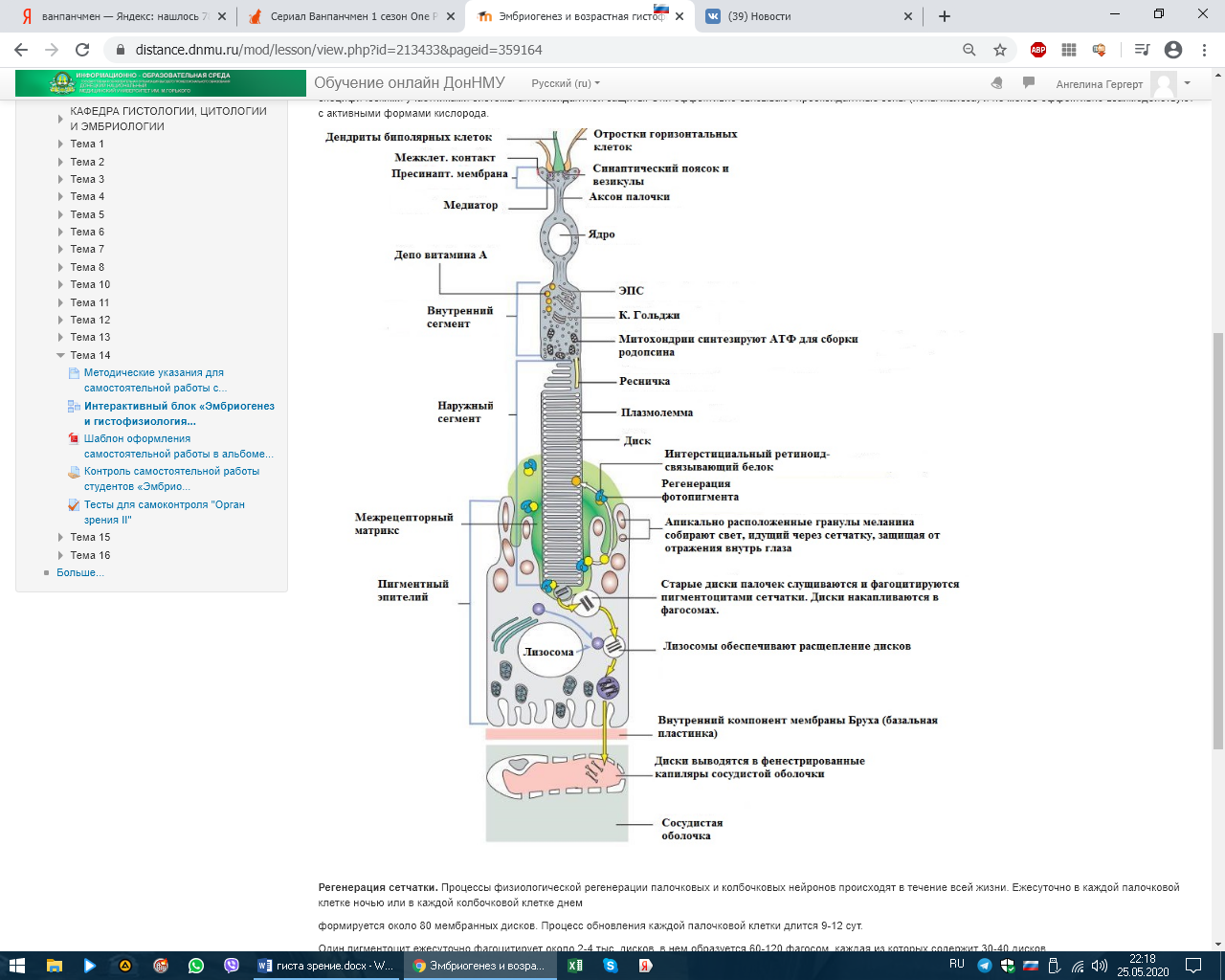 |