Сенсорные системы
 Скачать 1.12 Mb. Скачать 1.12 Mb.
|

|
Локомоция (управление движением)
Функции спинного мозга Спинной мозг выполняет две основные функции: рефлекторную и проводниковую. 1.рефлекторная — осуществляется соматической и вегетативной нервными системами. Заключается в том, что его двигательные нейроны (мотонейроны) управляют движениями мышц конечностей, туловища и отчасти шеи. Через спинной мозг проходят рефлекторные дуги, с которыми связаны сокращения всех скелетных мышц тела (кроме мышц головы). 2.проводниковая —Заключается в том, что по волокнам белого вещества информация от кожных рецепторов (прикосновения, боли, температурных), рецепторов мышц конечностей и туловища, рецепторов сосудов, органов мочеполовой системы передаётся по восходящим проводящим путям в головной мозг. И наоборот, от двигательных центров головного мозга импульсы поступают к мотонейронам передних рогов, а оттуда – к органам (к мышцам конечностей, туловища и т.д.).  Спинномозговые рефлексы являются врожденными, их осуществление не требует осознания. Сухожильные спинальные рефлексы являются клинически важными, т.к. каждый из них замыкается в определенных сегментах спинного мозга. Поэтому, по характеру рефлекторной реакции можно судить о функциональном состоянии соответствующих сегментов спинного мозга. Клетки Реншоу (КР)это тормозные вставочные нейроны, расположенные в передних рогах спинного мозга, несколько дорзальнее и медиальнее, чем мотонейроны (МН)
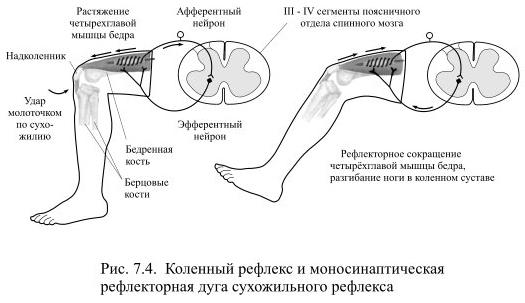 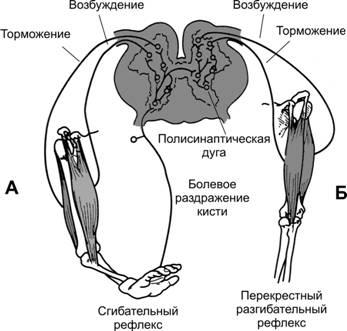 Рис. 13–7. Реципрокное торможение. А — сгибательный рефлекс. Б — перекрестный разгибательный рефлекс. Сгибательный рефлекс возникает в ответ на сильные раздражения болевых рецепторов конечности и заключается в сокращении сгибательных мышц с одновременным расслаблением разгибательных мышц, чтобы удалить конечность от источника раздражения (рис. 13–7). Болевое раздражение, наносимое на какой-либо участок тела, также вызывает реакцию ухода от раздражения и носит более общее название — рефлекс отдёргивания.
Концепция Н. А. Бернштейна (1947) о построении движений имела огромное значение для создания теории произвольного двигательного акта. Согласно данной концепции, любое движение — сложная многоуровневая система, где каждый уровень (или определенные анатомические структуры) характеризуется «ведущей афферентацией» и собственным набором регулируемых движений. Н. А. Бернштейном выделены пять уровней регуляции движений: Уровень А – руброспинальный – субкортикальный уровень по положению, в его обеспечении участвуют подкорковые образования. Ведущая афферентация – проприорецептивная чувствительность, но не вся, а та часть, которая указывает на напряжение (тонус) мышц. Этот уровень обеспечивает двигательные действия типа дрожи (изменения тонуса мышц), обмахивания (напряжение/расслабление мышц), напряжение тела (напряжение мышц для поддержания определённой позы). Основная функция – создание фона для других. Нарушение уровня:А)дистонии (нарушение тонуса мышц) по двум типам: гипо- и гипертонус – гипертония (каталепсия) – чрезмерное напряжение мышц, гипотонии – восковая гибкость. Паркинсонизм – нарушение тонуса мышц. Б)тремор – дрожание различных частей тела в покое или движении. Уровень В – таламополидарный уровень – уровень синергий и штампов. Подкорковый уровень, ведущими афферентами является проприорецептивная информация – суставно-угловая чувствительность, тот вид чувствительности, который даёт информацию о том, как разные части тела расположены относительно друг друга. Этот уровень обеспечивает обширные мышечные синергии - содружественные движения, которые носят постоянный неменяющийся характер (походка). Нарушения протекают по типу гипо и гиперфункции. Гипо – выпадение движений из слаженного ансамбля, гипер – включение лишних движений, нарушения – диссинергии. Примером по типу выпадения является паркинсонизм (скованность позы и др.), по типу лишних движений – различного рода сложные движения патологического характера, когда движения недостаточны для достижения цели, но с включением дополнительных. Уровень С – пирамидно-стриальный уровень – уровень пространственного поля. Кортикальный уровень, ведущая афферентация – синтетическое пространственное поле (пространственная информация носит разномодальный характер). Движения на этом уровне осуществляются в пространственном поле и носят целевой характер. Для выполнения всегда необходимо конкретное место в пространстве. Сюда входят локомоции (перемещение в пространстве), движения с конечным эффектом (забить гвоздь). Нарушения – атоксии – это утеря целевого характера движения (симультанная агнозия, больной не способен воспринимать сразу несколько объектов). Уровень Д – теменно-премоторный уровень (морфологическая характеристика) – уровень действий (психическая характеристика). Котрикальный, ведущая афферентация – предмет, его смысловое содержание. Движения на этом уровне обеспечивают различные предметные действия, которые определяются функциональным назначением предмета. Нарушения приводят к апроксиям. Уровень Е – уровень смысловых условных действий, на котором формируется программа действия - связан с лобными долями, которые отвечают за программирование действий. При поражении – системные персеверации.
Децеребрационная ригидность – относительное расслабление мышц сгибателей и резкое повышение тонуса мышц разгибателей, происходящее вследствие децеребрации (перерезки ствола головного мозга). Положение туловища с опорой на затылок и пятки. (опистотонус). Происходит из-за высвобождения центров продолговатого и спинного мозга из под контроля ретикулярной формации продолговатого и среднего мозга. Продолговатый мозг отвечает за тонус скелетных мышц, его вестибулярные ядра за тонус разгибателей, неспецифические отделы за сгибатели. Продолговатый мозг участвует в организации установочных рефлексов тела. За распределение тонуса при поддержании или изменении позы отвечает рубро-спинальный путь, черная субстанция отвечает за поддержание точной позы. При перерезке ствола мозга ретикулярная формация перестает осуществлять регулирующий контроль.
Первичная моторная кора (прецентральные извилины) получает информацию от соматосенсорной коры м управляет двигательными центрами ствола и СМ контролирующих сокращение склеенной мускулатуры. Первичная моторная кора каждого полушария контролирует мышцы на противоположной стороне тела. Площадь поверхности областей первичной моторной коры, управляющих различными частями тела, пропорциональна не физической поверхности этих частей, а числу и степени сложности их моторных реакций. Поражения прецентральной извилины вызывают определённой группы мышц противоположной стороны тела. Вторичная моторная зона (поле 6) расположена как впереди прецентральной извилины (премоторная кора), так и на медиальной поверхности, соответствующей коре верхней лобной извилины. Вторичная двигательная кора осуществляет высшие двигательные функции, связанные с планированием и координацией произвольных движений. Раздражение коры поля 6 вызывает сложные координированные движения: поворот головы, глаз и туловища в противоположную сторону, содружественные сокращения сгибателей или разгибателей на противоположной стороне. В премоторной коре расположены двигательные центры, связанные с социальными функциями человека: центр письменной речи в заднем отделе средней лобной извилины (поле 6), центр моторной речи Брока, а также музыкальный моторный центр (поле 45). Нейроны двигательной коры получают афферентные входы через таламус от мышечных, суставных и кожных рецепторов, от базальных ганглиев и мозжечка. Основным эфферентным выходом двигательной коры на стволовые и спинальные моторные центры являются пирамидные клетки V слоя.
3 области, отличающиеся спецификой соединения со Сп.мозгом и головным и выполняемыми ими функциями: 1) Вестибулоцеребеллум — получает афф- инф-цию от вестибулярных ядер, посылает к ним эфф- сигналы. Задача: сохранение равновесия при стоянии и ходьбе, управление движениями глаз. 2) Спиноцеребеллум — получает инф-цию от афф- путей Сп.м: два спиномозжечковых тракта, несущие импульсы от проприорецепторов мышц и сухожилий. Эфф- инф-ция поступает к двигательным центрам ствола (мед. и лат. нисходящие пути). Функция: контроль правильности начинающихся движений рук и ног. 3) Цереброцеребеллум — получает при участии ядер моста инф-цию о планирующемся движении из сенсомоторных областей коры и посылает эфф- сигналы к первич. и вторич. мотороной коре. Участие в планировании движений. Мозжечок не может инициировать движение, он лишь обеспечивает необходимый баланс действующих мышц, особенно в заключительной стадии движений. При мозжечковом поражении пациент, например, промахивается, когда его просят быстро прикоснуться к кончику своего носа указательным пальцем.  Для проверки функций мозжечка в клинической практике применяется несколько проб, сущность которых сводится к выявлению имеющихся нарушений координации движений.
Проба Ромберга служит для проверки устойчивости испытуемого. Предложить испытуемому встать, плотно сдвинув стопы (голова чуть приподнята, руки опущены вдоль туловища). Экспериментатор при этом должен находиться сзади, страхуя испытуемого от падения. В случае удовлетворительного выполнения этой пробы позу усложняют, предложив испытуемому, не меняя положения ног, вытянуть вперед руки и закрыть глаза. Если равновесие сохраняется, то позу еще более усложняют, предлагая испытуемому установить стопы вдоль одной линии (руки вытянуты вперед, глаза закрыты). В последней позе и у здоровых субъектов может наблюдаться некоторая неустойчивость.
Проводятся с целью проверки возможности выполнения различных произвольных движений конечностями. а) Пальце-пальцевая проба. Предложить испытуемому сесть, закрыть глаза и развести руки в стороны. Из такого исходного положения он должен коснуться указательными пальцами друг друга, медленно сводя руки. б) Пальце-носовая проба. Предложить испытуемому сначала с открытыми, а затем с закрытыми глазами коснуться кончика носа указательным пальцем из положения выпрямленной и отведенной в сторону руки. При поражении мозжечка, выполняя пробы а) и б), испытуемый промахивается мимо цели. Также может проявляться дрожание руки в начале движения и при приближении к цели (интенционный тремор). в) Проба на адиадохокинез (последовательное совершение противоположных по характеру движений). Предложить испытуемому согнуть руку в локтевом суставе под прямым углом, пальцы разведены и слегка согнуты. В таком положении быстро совершать пронацию и супинацию (имитация вкручивания электрической лампочки). При поражении мозжечка движения замедленны, неловки и асинхронны. г) Проба на асинергию. Предложить испытуемому согнуть руку в локтевом суставе и удерживать ее согнутой около собственной груди, глаза закрыть. Экспериментатор, упираясь одной рукой в грудь испытуемого, пытается другой рукой разогнуть его предплечье, а затем неожиданно отпускает руку испытуемого. У здорового человека быстрое включение в действие мышц-антагонистов предотвращает удар в собственную грудь.
Рецепторы вестибулярного аппарата расположены в области преддверия полукружных каналов, залегающих в перепончатом лабиринте улитки. Рецепторный эпителий располагается в области маточки и мешочка – двух расширений области преддверия. Вследствие действия различных ускорений, отолитовы мембраны, на которых расположен рецепторный эпителий, смещается, что вызывает раздражение вторичных рецепторных клеток. Существуют также рецепторы, расположенные в ампулах полукружных каналов, они представляют собой эпителиальные клетки с множеством ворсинок, которые погружены в эндолимфу. При различных изменениях положения тела эндолимфа смещается, вызывая деформацию ворсинок эпителия, что является импульсом для вестибулярного анализатора. Таким образом за восприятие линейного ускорения отвечают маточка и мешочек, а за восприятие линейного – эпителий ампул полукружных каналов. Рецепторные эпителиальные клетки являются рецепторами вторичными, они с помощью синапсов соединяются с периферическими окончаниями нейронов вестибулярного ганглия. Рецепторные клетки имеют стереоцилии и киноцилии – волосковые отростки, при смещении стереоцилий в сторону киноцилии клетки деполяризуется, а при смещении их в противоположную сторону – гиперполяризуется. Таким образом клетка имеет два активных полюса и может либо возбуждаться, либо тормозиться. Плюс ко всему у рецепторов есть спонтанныя фоновая активность. Информация с аксонов первичных рецепторов передается на нейроны вестибулярных ядер предверно-улиткового нерва, далее направляются аксоны этих ядер в мосто-мозжечковый угол,где находятся ядра Дейтерса и Швальбе. Их аксоны формируют преддверно-таламический тракт и идут в таламус, часть аксонов переключается дальше и дет к коре (средней и нижней височным извилинам . Вестибулярные реакции обусловлены различными проводящими путями. Многие из них заканчиваются в спинном мозге и обуславливают перераспределение тонуса мышц при изменении положения тела, пути оканчивающиеся в коре позволяют осознавать положение тела в пространстве, а пути ассоциированные с гипоталамусом обуславливают появление кинетозов, например морской болезни.
1) Специфические/релейные ядра: Сенсорные:
Чувствительные ядра передают информацию в первичную сенсорную кору. Моторные:
Лимбические – передние. 2) Неспецифические ядра модулируют активность больших областей коры. К ним относятся ядра средней линии, внутрипластинчатые ядра и ретикулярное ядро таламуса. 3) Ассоциативные ядра - полимодальные связаны с ассоциативными областями коры. К ассоциативным ядрам относятся латеральное дорзальное, латеральное заднее, дорзомедиальное ядро, ядра подушки зрительного бугра.
К стириопаллидарной системе (базальным ядрам) относят: хвостатое ядро+скрлупа= полосатое тело, бледный шар и субталамическое ядро. С системой тесно связана sustantia nigra (SNr) среднего мозга, влияющая на передачу сигналов от полосатого тела к внутренней и наружной частям бледного шара. Схема их взаимодейсвия на картинке. В SNr нейроны производят дофами. Ч/з D1 рецепторы усиливается торможение, ч/з D2 ослабляется торможение. Активация премоторной и моторнной лобной области коры → возбуждение полосатого тела, в кот. преобладают эфф- тормлзные ГАМКэогич. нейроны → уменьшают спонтанную активность внутренней части бледного шара и SNr. Меньшая часть выходных нейронов полосат.тела используют медиатор в-во Р — возбуждающ. + многие нейроны полосат.тела синтезирую энкефалины — нейромодуляторы, и дейсвие оказывается торможно-модулирующим. От бледного шара и SNr сигналы передаются вентролатеральным ядерам таламуса → проекция на премоторные области коры. Субталамическое ядро получает топографически организованные сигналы от премоторной и моторной коры, в соот. с которыми оно регулирует выходную активность бледного шара. 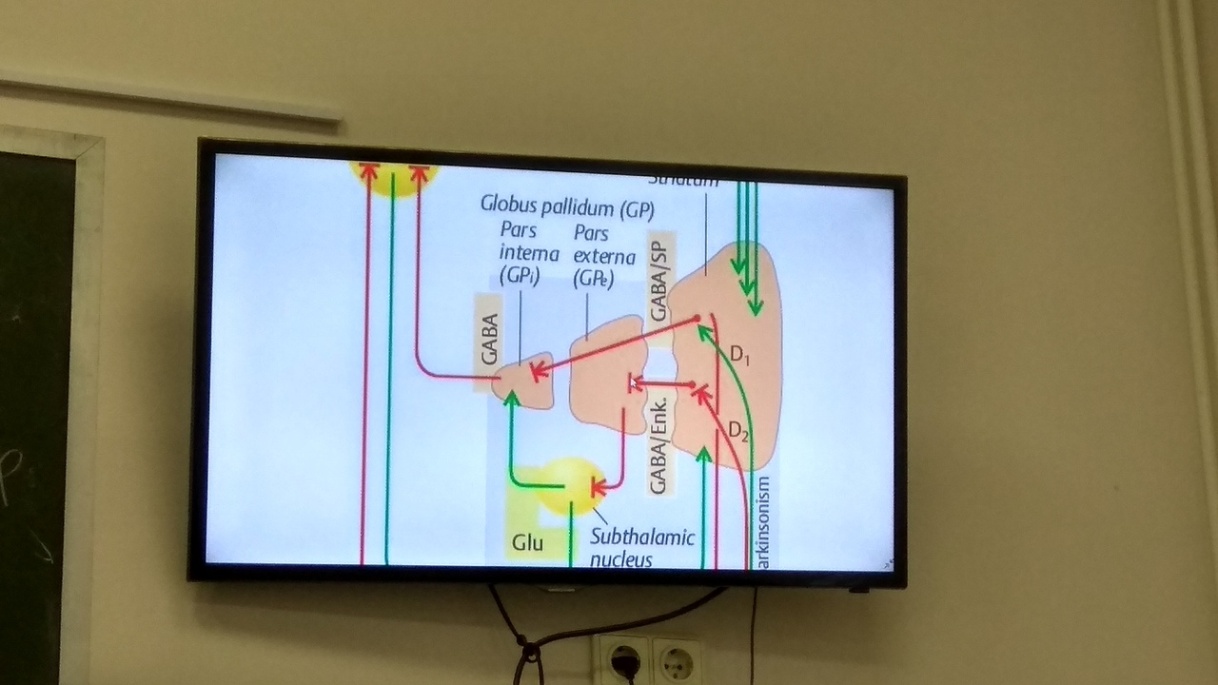 Конечным результатом является создание в моторной коре двигательных программ, предназначенных для исолнения двигательными центрами ствола и нейронами Сп.мозга. Функция: участие в планировании и инициации произвольных движений; участие в познават.деят-ти мозга, в формировании эмоций.
Надсегментарный - стволовые структуры ГМ - двигательные ядра продолговатого и среднего мозга, отделы ретикулярной формации и мозжечка. Высшие отделы головного мозга осуществляют свои управляющие функции че рез две нисходящие системы — экстрапирамидную и пира мидную. Экстрапирамидная система - участвует в организации движений, мышечного тонуса и позы посредством нисходящих двигатель ных путей. Функция заключается в осуществлении позно-тонических реак ций, циклических движений, удержании равновесия тела и взаимной координации позных рефлексов и целенаправленного движения, в коррек ции выполняющихся движений. Экстрапирамидная система оказывает свое влияние на мо тонейроны спинного мозга посредством трех нисходящих пу тей. Это кортико-рубро-спинальный путь, в который включены красное ядро, мозжечок, промежуточный мозг, базальные ган глии. Этим путем осуществляется организация позно-тониче ских реакций. Второй путь — кортико-ретикуло-спинальный, име ющий основную функцию — тоническое повышение возбуди мости мотонейронов сгибателей. Третий — кортико-вестибуло-спинальный, посредством которого обеспечивается актива ция мотонейронов мышц-разгибателей. Пирамидная система - это система управления движением включающая в себя преимущественно двига тельные зоны больших полушарий и сегментарные двига тельные нейронные структуры. Включает нисходящие пути двигательной коры к мо торным ядрам черепномозговых нервов. В пирамидный тракт посылают свои аксоны пирамидные нейроны зрительной, слуховой, соматосенсорной, ассоциативной зон коры. Аксоны этих нейронов, не преры ваясь, спускаются по соответствующей половине мозгового ствола до уровня пирамид продолговатого мозга. На этом уровне осуществля ется переход до 80-90 % всех волокон на противоположную сторону и, постепенно истончаясь, пирамидный тракт закан чивается в нижних сегментах спинного мозга на вставочных и двигательных его нейронах. По прохождении через ствол мозга часть волокон дает коллатерали к ядрам ствола (крас ное ядро, ретикулярная формация, сенсорные ядра череп номозговых нервов). Оставшиеся 10-20 % не перекрещенных волокон переходят на противоположную сторону уже на уров не сегментов, где они заканчиваются. В состав пирамидного тракта входят: · Αδ-волокна — тонкие немиелинизированные. Аксоны мелких и средних пирамидных нейронов. · Толстые, миелинизированные. Принадлежат крупным пирамидным клеткам Беца. · Быстрые - толстые волокна, имею щие скорость распространения возбуждения в пределах 60 -120м/с. · Медленные -1-20 м/с. Это позво лило предположить наличие в пирамидной системе двух ме ханизмов управления движением: быстрого, для выполнения срочных фазических двигательных актов, и медленного, регу лирующего позные реакции. Пирамидные влияния приводит к торможению мотонейронов, участвующих в тонических антигравитационных рефлексах, и к активации мотонейронов, реализующих фаз ные сгибательные рефлексы. Пирамидной системе принадлежит важная роль в реа лизации быстрых произвольных движений, осуществляе мых под контролем сознания. Эта функция реализуется в тесной связи с экстрапирамидной системой. В целом функция пирамидной сис темы сводится к контролю сформированной в головном мозге программы двигательного акта. Тонические рефлексы регулируют положение тела. Дуга их замыкается в стволе мозга. Различают шейные и лабиринтные тонические рефлексы. Шейные возникают в ответ на раздражение, связанное с изменением положения головы по отношению к туловищу, а лабиринтные — вызываются раздражениями, идущими из перепончатых лабиринтов в связи с перемещением головы в пространстве. 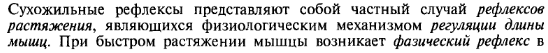  |