Сенсорные системы
 Скачать 1.12 Mb. Скачать 1.12 Mb.
|

|
Сенсорные системы
Сенсорные системы = орган чувств (правда сохранился только для обозначения анатомически обособленных периферич отделов некоторых сенсорных систем (глаз,ухо). Все сенсорные системы состоят из периферических рецепторов, проводящих путей и переключательных ядер, первичных проекционных областей коры и вторичной сенсорной коры. Рецепторы обладают специфичностью, т. е. высокой возбудимостью только к определенным раздражителям, получившим название адекватных. В частности, для глаза адекватным раздражителем являются световые, а для уха – звуковые волны и т. д. При действии адекватных раздражителей возникают ощущения, характерные для определенного органа чувств. Так, раздражение глаза вызывает зрительные ощущения, уха – слуховые и т. д. Кроме адекватных, существуют и неадекватные раздражители, которые вызывают только незначительную часть ощущений, свойственных данному органу чувств, или действуют необычным образом. Например, механическое или электрическое раздражение глаза воспринимается как яркая вспышка света («фосфен»), но не дает образа предмета и восприятия цветов. Специфичность органов чувств является результатом приспособления организма к условиям внешней среды. Адекватные раздражители- стимулы, к которым рецепторы наиболее чувствительны.  Принцип меченой линиизаключается в пространственно упорядоченном расположении нейронов на различных уровнях сенсорных систем соответственно характеристикам их рецептивных полей. Цепь модально-специфичных нейронов, соединенных синапсами в определенную жесткую цепь, передающую информацию только от своего рецептивного поля. 
Рефракция глаза – свободное преломление световых лучей в глазу, без аккомодации. Или преломляющая способность оптической системы глаза. К аномалиям рефракции относят:
Способы коррекции : очки (искусственное изменение степени преломления лучей) при близорукости линзы двояковогнутые, а при дальнозоркости – двояковыпуклые, при астигматизме величина выпуклости линз меняется по осям. Также возможна лазерная коррекция зрения, основанная на прижигании роговицы и изменении ее толщины, применяют при близорукости, дальнозоркости и астигматизме.
Это области кожи с повышенной болевой чувствительностью (дерматомы), болезненность которых обусловлена заболеваниями внутренних органов (спланхотомы). На определенных участках кожи проявляется так называемая отраженная боль, а также повышение температуры и покраснение. Анатомически наличие таких зон связано с сегментарным строением сп инного мозга. Если внутренний орган страдает, импульсы от патологии иррадиируют по чувствительному волокну в отдел СМ, это вызывает локальное возбуждение этого сегмента. В нем включается вторая связка нейронов – часть обеспечивает чувствительность кожи, и часть – эфферентные. При длительном возбуждении нервных центров СМ происходит истощение как афферентных, так и эфферентных нейронов, что приводит к патологическим изменениям кожных областей. По СМ патологический импульс проходит в ГМ, что также формирует устойчивое болевое ощущение соответствующей кожной зоне.
схема по тексту Ткаченко. 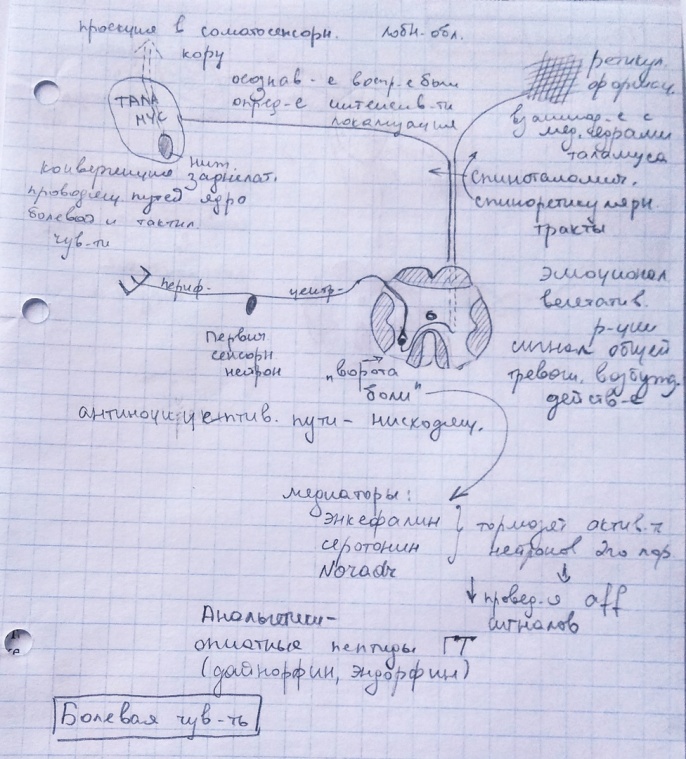 Боль определяется как неприятное сенсорное ощущение и эмоциональное переживание, связанная с повреждением ткани. Всегда субъективно неприятна и служит сигналом повреждения или болезни, побуждает к прекращению контакта с повреждающими факторами окр.среды. Ноцирецепторы (болевые) - свободные нервные окончания, расположенные в коже, слизистых обол., мышцах, суставах, надкостнице, внутр.органов. Различие ранней боли (короткой и острой) происходит по миелинизировннам волокнам (высок скорость передачи импульса), и поздней боли )тупой, длительной) — по безмякотным волокнам. Ноцирец-ры полимодальны, активируются стимулами разной природы: химич, механич., термич. Порог чувс-ти высок. Схема пути боли на фото. 2 пути от «ворот боли» - синапс с нейронами второго порядка в задних рогах Сп.м.: 1) Спиноталамический (через нейроны нижнего заднелатерального ядра таламуса, далее проекция на соматосенсорную кору) — обеспечивает осознаваемое воспр-ие боли, определение интенсивности стимула, его локализация. 2)Спиноретикулярный ( конец на нейронах ретикуляр.формации, взаимодействующих с медиальными ядрами таламуса) — роль сигнала общей тревоги, оказывает на человека общее возбуждающее дей-е, сопровождение эмоциональными и вегетативными реакциями. Антиноцицептивная система выполняет функцию «ограничителя» болевого возбуждения. Эта функция заключается в контроле за активностью ноцицептивных систем и предотвращении их перевозбуждения. Проявляется ограничительная функция в увеличении тормозного влияния антиноцицептивной системы в ответ на нарастающий по силе ноцицептивный стимул. Однако это ограничение имеет предел и при активность нейронов на ноцицептивную импульсацию, формируя у людей состояние анальгезии. Одновременно эндорфины активизируют антиноцицептивную систему. Субъективную оценку боли опредет соотношение нейронной активности обоих путей и зависимая от неё активация антиноцицептивных нисходящ. путей, способных изменить хар-тер проведения сигналов от ноцирецепторов. Сущ-ет эндогенный мех-зм уменьшения болевой чувст-ти путём регуляции порога синаптич. переключений в задних рогах Сп.мозга («ворота боли»). В них на передачу возб-ния влияют нисходящие волокна нейронов серого в-ва водопровода, голубого пятна, нек. ядер срединного шва. Медиаторы: энкефалин, серотонин, Noradr, тормозят активной нейронов второго порядка в зад.рогах Сп.мозга, чем уменьшают проведение афф-ных сигналов от ноцирец-ров. Опиатные пептиды (синез. Гипоталамуса) присоединяются к специфич. рец-рам нейронов лимбич. Системы и медальной области таламуса и активируют антиноцициептив. мех-мы и повышается порог болей чувств-ти в результате их повышенного образования. Баланс м/у ощущением боли и её субъектив.оценкой устанавливается с помощью лобных обл.мозга.
Источником возбуждений, поддерживающих мышечный тонус, являются проприорецепторы. В скелетных мышцах имеются три вида проприорецепторов: • мышечные веретена, расположенные среди мышечных волокон; • сухожильные рецепторы Гольджи, расположенные в сухожилиях; • пачиниевы тельца, расположенные в фасциях, сухожилиях, связках. Особое значение в регуляции мышечного тонуса имеют мышечные веретена и сухожильные рецепторы Гольджи. Мышечные веретена представляют собой небольшие продолговатые образования, напоминающие своим внешним видом прядильные капсулы мышечного веретена находится пучок мышечных волокон, которые называются интрафузальными, т. к. они расположенными внутри веретена в отличие от обычных мышечных волокон, которые называются зкстрафузальными. Каждое интрафузальное волокно состоит из трех частей: • его центральная часть называется ядерной сумкой, в которой находятся ядра мышечной клетки; • два периферических участка, которые имеют поперечную исчерченность и обладают способностью сокращаться; • миотрубки, расположенные между ядерной сумкой и периферическими участками. Ядерную сумку в виде спирали окружают нервные волокна чувствительного нейрона-первичные рецепторные окончания. В области миотрубок нервные окончания афферентных нейронов гроздевидно ветвятся, образуя вторичные рецепторные окончания. В мышце мышечное веретено одним концом прикрепляется к экстрафузальному мышечному волокну, а другим - к сухожилию этого волокна. Таким образом, мышечное веретено расположено в мышце параллельно экстрафузальным мышечным волокнам. При снижении тонуса экстрафузального волокна увеличивается его длина, что приводит к растяжению и раздражению первичных и вторичных рецепторных окончаний, для которых растяжение является адекватным раздражителем. Моторные отделы нервной системы координируют работу мышц путем управления активностью соответствующих мотонейронов спинного мозга. И хотя регуляция движений отчасти произвольна, она осуществляется прежде всего посредством рефлекторных актов и механизмов, не связанных с сознанием. Важную роль играют спинномозговые рефлексы. По типу адекватного раздражителя мышечные веретена и сухожильные органы относятся к рецепторам растяжения. Однако их расположение различно; первые лежат параллельно экстрафузальным волокнам, а вторые соединены с ними последовательно. В результате неодинаков и характер их активации. Во время расслабления (растяжения) мышцы активность мышечных проприорецепторов возрастает, при сокращении же мышечных волокон напряжение мышечных веретен снижается и импульсация от них прекращается. Однако сухожилия при этом растянуты и частота разрядов сухожильных рецепторов увеличивается. Следовательно, мышечные веретена регистрируют главным образом увеличение длины мышцы, а сухожильные органы – увеличение ее напряжения. 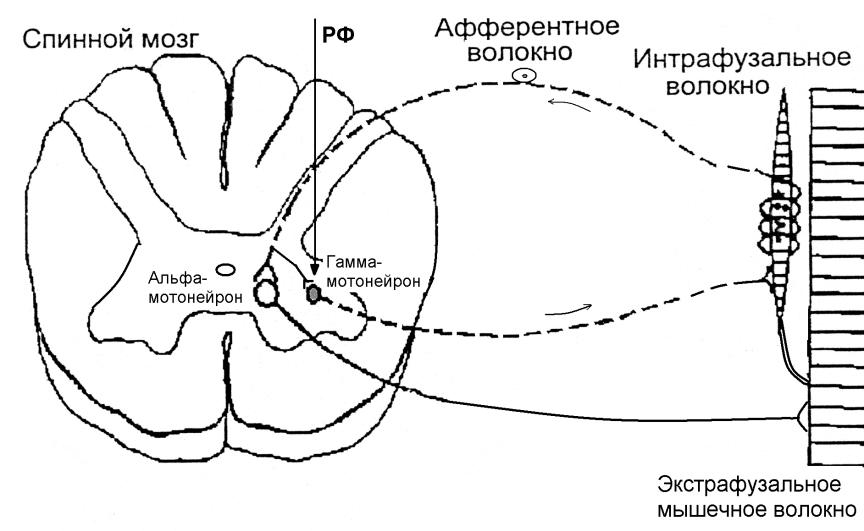 Периферические мотонейроны подразделяются на альфа-мотонейроны и гамма-мотонейроны (рис. 21.2 ). Меньшие по размеру гамма-мотонейроны иннервируют интрафузальные мышечные волокна . Активация гамма-мотонейронов увеличивает растяжение мышечных веретен, тем самым облегчаясухожильные и другие рефлексы, замыкающиеся через альфа-мотонейроны. Каждую мышцу иннервирует несколько сотен альфа-мотонейронов. В свою очередь, каждый альфа-мотонейрон иннервирует множество мышечных волокон - около двадцати в наружных мышцах глаза и сотни в мышцах конечностей и туловища . В нервно-мышечных синапсах выделяется ацетилхолин . Аксоны периферических мотонейронов идут в составе черепных нервов и передних корешков спинного мозга . На уровне межпозвоночных отверстий передние корешки и задние корешкисливаются, образуя спинномозговые нервы . Несколько соседних спинномозговых нервов образуют сплетение, а затем разветвляются на периферические нервы . Последние тоже неоднократно разветвляются и иннервируют несколько мышц. Наконец, аксон каждого альфа-мотонейрона образует многочисленные разветвления, иннервируя многие мышечные волокна. Каждый альфа-мотонейрон получает прямые возбуждающие глутаматергические входы от корковых мотонейронов и от чувствительных нейронов , иннервирующих мышечные веретена . Возбуждающие влияния поступают также к альфа- и гамма-мотонейронам от двигательных ядер ствола мозга и вставочных нейронов спинного мозга - как по прямым путям, так и с переключениями. Прямое постсинаптическое торможение альфа-мотонейронов осуществляют клетки Реншоу - вставочные глицинергические нейроны . Непрямое пресинаптическое торможение альфа-мотонейронов и непрямое пресинаптическое торможение гамма-мотонейронов обеспечивают другие нейроны , образующие ГАМКергические синапсы на нейронах задних рогов . Тормозное действие на альфа- и гамма-мотонейроны оказывают и другие вставочные нейроны спинного мозга, а также двигательные ядра ствола мозга . Если преобладают возбуждающие входы, группа периферических мотонейронов активируется. Вначале возбуждаются мелкие мотонейроны. По мере того как сила сокращения мышцы нарастает, частота их разрядов нарастает и вовлекаются крупные мотонейроны. При максимальном сокращении мышцы возбуждена вся соответствующая группа мотонейронов. |