зобез. Система протистов
 Скачать 2.33 Mb. Скачать 2.33 Mb.
|

|
Гипогенез – жизненные циклы у полипов. 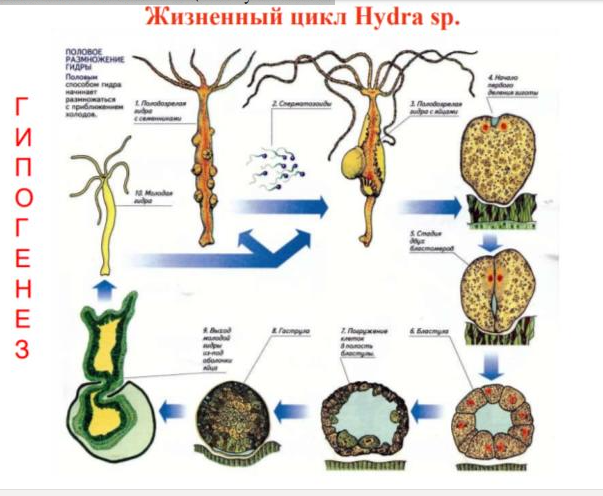 14. Пресноводная гидра. Строение. Клеточный состав. Питание. Размножение; Строение: Гидра (Hydra) — типичный представитель кишечнополостных животных. Форма ее тела трубковидная, на переднем конце находится ротовое отверстие, окруженное венчиком из 5—12 щупалец. Сразу под щупальцами у гидры имеется небольшое сужение — шейка, отделяющая голову от туловища. Задний конец гидры сужен в более или менее длинную ножку, или стебелек, с подошвой на конце. Сытая гидра имеет длину не более 5—8 миллиметров, голодная значительно длиннее. Тело гидры, как и у всех кишечнополостных, состоит из двух слоев клеток. Во внешнем слое клетки разнообразны: часть из них выполняют роль органов поражения добычи (стрекательные клетки), другие выделяют слизь, третьи обладают сократимостью. Во внешнем слое рассеяны и нервные клетки, отростки которых образуют сеть, покрывающую все тело гидры. Гидра — один из немногих представителей пресноводных кишечнополостных, основная масса которых является обитателями моря. В природе гидры встречаются в различных водоемах: в прудах и озерах среди водных растений, на корешках ряски, зеленым ковром покрывающей канавы и ямы с водой, небольшие пруды и речные заводи. В водоемах с чистой водой гидр можно встретить на голых камнях у самого берега, где иногда они образуют бархатистый ковер. Гидры светолюбивы, поэтому держатся обычно на неглубоких местах вблизи берегов. Они способны различать направление потока света и двигаются в сторону его источника. При содержании в аквариуме всегда перебираются на освещенную стенку. Если набрать в сосуд с водой побольше водных растений, то можно наблюдать гидр, ползающих по стенкам сосуда и листьям растений. Подошва гидры выделяет клейкое вещество, за счет которого она прочно закрепляется на камнях, растениях или стенках аквариума, и отделить ее нелегко. Изредка гидра перемещается в поисках пищи. В аквариуме можно отмечать ежедневно точкой на стекле места ее прикрепления. Такой опыт показывает, что за несколько дней передвижение гидры не превышает 2—3 сантиметров. Чтобы переменить место, гидра временно приклеивается к стеклу щупальцами, отделяет подошву и подтягивает ее к переднему концу. Прикрепившись подошвой, гидра распрямляется и снова опирается щупальцами еще на шаг вперед. Такой способ передвижения похож на то, как ходит гусеница бабочек пядениц, называемая в просторечьи «землемер». Только гусеница подтягивает задний конец к переднему, а затем снова двигает вперед головной конец. Гидра же при такой ходьбе все время переворачивается через голову и таким образом передвигается относительно быстро. Существует и другой, гораздо более медленный способ передвижения — скольжение на подошве. Усилием мускулатуры подошвы гидра еле заметно сдвигается с места. Некоторое время гидры могут плавать в воде: открепившись от субстрата, расправив щупальца, они медленно падают на дно. На подошве может образовываться пузырек газа, который увлекает животное вверх. Питание Гидра — хищник, пищей ей служат инфузории, мелкие рачки — дафнии, циклопы и другие, иногда попадается более крупная добыча в виде личинки комара или небольшого червя. Рыбоводческим прудам гидры даже могут наносить вред, поедая выклюнувшихся из икры мальков рыб. Охоту гидры легко наблюдать в аквариуме. Широко раскинув свои щупальца, так что они образуют ловчую сеть, гидра висит щупальцами вниз. Если долго наблюдать за сидящей гидрой, можно видеть, что ее тело все время медленно раскачивается, описывая передним концом круг. Плывущий мимо циклоп задевает за щупальца и начинает биться, чтобы освободиться, но скоро, пораженный стрекательными клетками, затихает. Парализованная добыча подтягивается щупальцем ко рту и поглощается. При успешной охоте маленький хищник раздувается от проглоченных рачков, темные глазки которых просвечивают сквозь стенки тела. Гидра может проглотить и добычу крупнее ее самой. При этом рот хищницы широко раскрывается, а стенки тела растягиваются. Иногда изо рта гидры торчит часть непоместившейся добычи. Размножение При хорошем питании гидра быстро начинает почковаться. Рост почки от маленького бугорка до вполне сформированной, но еще сидящей на теле материнской особи, гидры занимает несколько дней. Часто, пока молодая гидра еще не отделилась от старой особи, на теле последней уже образуется вторая и третья почки. Так происходит бесполое размножение, половое размножение наблюдается чаще под осень при понижении температуры воды. На теле гидры появляются вздутия — половые железы, одни из которых содержат яйцевые клетки, а другие — мужские половые клетки, которые, свободно плавая в воде, проникают в полость тела других гидр и оплодотворяют неподвижные яйцеклетки. После образования яиц старая гидра обычно погибает, а из яиц при благоприятных условиях выходят молодые гидры. 15. Строение и жизненные циклы сцифоидных медуз; Внешне сцифоидные медузы имеют значительное сходство с гидроидными, но значительно превышают их по размерам. Форма колокола, его расцветка и бахромчатые края, ротовые лопасти и многочисленные щупальца сцифоидных медуз делают их самыми красивыми организмами, обитающими в море. Тело сцифомедузы имеет форму диска, зонтика или колокола. В центре нижней стороны расположено ротовое отверстие, окруженное 4 ротовыми лопастями. В промежутках между ротовыми лопастями у многих сцифомедуз имеются впячивания, так называемые субгенитальные ямки. Они расположены под половыми железами и служат, по-видимому, для того, чтобы облегчить доступ кислорода к развивающимся половым продуктам. Край зонтика сцифомедузы имеет розетковидную форму, так как расчленен на 8 или 16 краевых лопастей, к их нижней стороне прикрепляются щупальца, а между ними находятся ропалии — маленькие, видоизмененные щупальца, несущие органы чувств — глазки и статоцисты. Число щупалец у разных групп сцифомедуз различно и колеблется от 4 до нескольких сотен. У корнеротых медуз щупальца отсутствуют. Гастральная полость сцифоидных медуз устроена довольно сложно. Рот ведет в короткую трубковидную глотку, которая открывается в желудок, имеющий 4 боковых кармана. Внутри карманов стенки желудка образуют многочисленные длинные энтодермальные выросты, так называемые гастральные нити. Они выделяют в кишечную полость пищеварительные соки. От желудка к периферии зонтика отходят радиальные каналы, число которых обычно равно или кратно 8. У ряда видов радиальные каналы ветвятся или даже образуют сеть. На краю зонтика радиальные каналы впадают в кольцевой, но иногда последний отсутствует. Нервная система сцифомедуз представляет собой сплетение нервных клеток. По краю колокола идет нервное кольцо, а вблизи ропалиев концентрируются нервные элементы, образующие здесь рыхлые скопления, сходные с ганглиями. Ропалий представляет собой маленькое недоразвитое видоизмененное щупальце, несущее в своей концевой части группу известковых телец — статолитов. Прикосновение кончика ропалия к чувствительным клеткам краевой лопасти вызывает раздражение, передающееся через ближайшее скопление нервных клеток мускулатуре зонтика, вызывая ее сокращение. Таким образом регулируется темп пульсации зонтика. У экземпляров с удаленными ропалиями регулярность сокращений нарушается. Ропалий, также как гидростатический орган, дает возможность медузе ориентировать тело ротовым отверстием вниз. У многих сцифоидных медуз на ропалии располагаются также светочувствительные органы — глазные пятна и более сложно устроенные глазки, напоминающие по строению описанные выше глазки гидромедуз. Сцифоидные медузы раздельнополы, их половые железы развиваются в энтодерме и располагаются в карманах желудка. Половые продукты выводятся наружу через ротовое отверстие. Мезоглея сцифоидных медуз, так же как и у гидромедуз, развита очень сильно. 16. Строение коралловых полипов. Геологическое значение; Тело полипа имеет такую структуру: эктодерма (наружный покров и поверхность глотки), мезодерма (гелеобразное вещество, наполняющее пустоты), и эндодерма (из нее сделаны внутренние стенки тела особи). Как мы уже говорили, у этих многоклеточных есть скелет. Причем, располагаться он может как снаружи, так и внутри. Что касается его состава, то это известь, либо рогоподобное вещество. Отметим, что коралловые полипы строение имеют схожее с гидроидными. Но никогда не переходят в стадию медузы. Само тело похоже на слегка деформированный цилиндр, на верхней части которого раскинут веер из щупальцев. В каждом таком «пальце» находятся особые капсулы, внутри которых заключена ядовитая субстанция. Умение ей пользоваться у кишечнополостных называется стрекательной функцией. Каждая такая опасная клетка имеет чувствительную ресничку. Если к полипу приблизилась жертва, либо он почуял опасность, и даже просто изменение давления воды, капсула раскрывается, из нее выскакивает стрекательная нить (трубка, сжатая спиралью в спокойном состоянии, по ней то и подается яд). Она впивается в тело жертвы, а токсичный секрет вызывает паралич и ожог тканей оппонента. После книдоцит (клетка) погибает, на его замену уже через двое суток приходит новый. Между щупальцами находится рот. Когда в него попадает что-то съедобное, сразу же отправляется в желудок через глотку. Она достаточно длинная и имеет форму сплюснутой трубочки. Весь этот коридор покрыт ресничками, создающими непрерывное движение тока воды внутри полипа. Благодаря чему животное получает, во-первых, еду (мелкий планктон), а во-вторых, дышит. Ведь обогащенная кислородом вода попадает внутрь его тела, а выводится уже насыщенная углекислым газом. Глотка заканчивается закрытой кишечной полостью. Она поделена на несколько отсеков. У основания кишечнополостные коралловые полипы расширяются. Если это одиночка, то такая база служит ему для того, чтобы прочнее прикрепиться к субстрату. Если же речь о колонии, то каждый ее член буквально врастает в общее с собратьями «тело» своим основанием. Как правило в одной системе состоят идентичные особи. Но существуют и такие колонии, где объединились разные полипы. Получение извести Колонии тропических коралловых полипов, разрастаясь, образуют барьерные рифы и коралловые острова – атоллы. Шестилучевые кораллы, составляющие основу рифов, благодаря известковому скелету широко используются для получения строительных материалов. Красные и чёрные скелеты восьмилучевых кораллов из Средиземного моря высоко ценятся в качестве украшений. 17. Общая характеристика Билатеральных (Трехслойных). Паренхиматозные, первичнополостные, целомические; Основные черты билатеральных: - В отличие от радиальных обладают двусторонней симметрией и трехслойностью строения , их тело развивается не из двух, а из трех зародышевых листков: эктодермы, энтодермы и мезодермы - У наиболее примитивных билатеральных животных рот расположен ближе к заднему концу тела, а у более прогрессивных групп смещен на передний конец, что дает преимущество в захвате добычи при движении головным концом вперед; - различают спинную, брюшную и боковую поверхности. - через их тело можно провести лишь одну (сагиттальную)плоскость симметрии; Билатеральные образуют два крупных подраздела: Бесполостные (acoelomata) и полостные животные(coelomata). Бесполостные ,или низшие черви включает несколько типов и характеризуется паренхиматозностью и отсутствием вторичной полости тела, выстланной мезодермальным эпителием.Ко вторичнополостным относится множество типов, в том числе высшие черви – кольчатые.Для них характерно наличие целома. Тело трехслойных животных (Triploblastica) в процессе онтогенеза формируется из трех зародышевых листков: экто-, мезо- и энтодермы. Особенностью строения представителей разных таксонов, относимых к данному уровню организации, является компактное расположение производных мезодермы в теле без формирования обширных полостей. Основной объем тела, заключенный между двумя эпителиями (экто- и энтодермой) заполнен телами клеток, их отростками и внеклеточным матриксом. Согласно классическим представлениям, сформировавшимся к середине ХХ века, весь этот комплекс получил название «паренхима». Однако такой ткани как «паренхима» не существует, и более корректным является использование термина «паренхиматозный тип организации». Планы строения паренхиматозных животных весьма сходны, вероятно, это определяется необходимостью иметь плоское тело, замкнутый кишечник и разветвленную протонефридиалную выделительную систему: отсутствие специализированной распределительной системы накладывает ограничения на размеры тела; замкнутая разветвленная кишка до определенной степени выполняет транспортную функцию; газообмен осуществляется всей поверхностью тела. Расположение ветвей кишечника в некоторой степени определяет характер расположения других систем органов в теле. “Plathelminthes” Плоские черви – не систематическая категория, это конструктивный тип организации, который характерен для трех таксонов типового ранга: Acoelomorpha, Catenulida и Rhabditophora. Плоские черви – билатерально-симметричные животные, тело которых уплощено в дорзо-вентральном направлении. Покровы представлены кожно-мускульным мешком, который представляет собой совокупность эктодермы (организованной в виде ресничного эпителия, либо тегумента), базальной пластинки и подстилающих ее мышечных элементов. В наиболее полном варианте состав мускулатуры кожно-мускульного мешка входят кольцевые, продольные, диагональные и дорсо-вентральные мышечные пучки. Степень развития мускулатуры коррелирует с размерами тела и образом жизни конкретных представителей. Пищеварительная система замкнутого типа имеется у большинства представителей; обычно она состоит из двух отделов: эктодермальной передней и энтодермальной средней кишки. Средняя кишка представлена либо мешковидным образованием, либо разветвленной конструкцией с главными ветвями и боковыми дивертикулами. Последнее характерно для крупных форм, у которых кишечник берет на себя выполнение транспортной функции. Выделительная система обычно протонефридиального типа: циртоциты более или менее равномерно распределены по всему телу животного, от них отходят собирательные капилляры, объединяющиеся в главные собирательные каналы. Собирательные каналы либо открываются наружу самостоятельными экскреторными порами, либо впадают в мочевой пузырек, который выделительным отверстием сообщается с окружающей средой. Специализированные транспортная и дыхательная системы отсутствуют. У аэробных форм газообмен осуществляется всей поверхностью тела. Нервная система ортогонального типа. На переднем конце тела располагается церебральный ганглий, от которого отходит несколько пар продольных нервных стволов, связанных между собой поперечными комиссурами. В некоторых случаях нервная система представлена плексусом. Половая система — гермафродитная, сложно устроенная, оплодотворение внутреннее. Для многих паразитических форм характерны сложные жизненные циклы с чередованием поколений и со сменой хозяев. На некоторых стадиях жизненного цикла у ряда представителей (у партенит трематод, ларвоцист цестод) формируется обширная первичная полость тела, которая выполняет транспортную и опорную функции. 18. Плоские черви: общая характеристика, классификация; 1. Билатерально-симметричные животные (Bilateria) 2. Тело формируется за счет трех зародышевых листков: экто- мезо- и энтодермы. 3. Уплощены в дорсовентральном направлении. 4. Наличие кожно-мускульного мешка. 5. Паренхиматозное строение (паренхима). 6. Кровеносная и дыхательная системы отсутствуют. Диффузия газов через поверхность тела. 7. Осморегуляторная и выделительная системы представлены протонефридиями. 8. Пищеварительная система состоит из двух отделов ― передней кишки и средней кишки. 9. Нервная система ортогонального типа (головной ганглий, несколько пар продольных нервных стволов, которые связаны между собой кольцевыми комиссурами). 10. Органы чувств: сенсиллы, глазки инвертированного типа, реже ― статоцисты. 11. Половая система ― гермафродитная. Внутреннее оплодотворение. 12. У подавляющего большинства Turbellaria развитие прямое (кроме Polycladida – Мюллеровская личинка). Все Neodermata развиваются с метаморфозом. 13. Для некоторых ресничных червей характерно бесполое размножение. (простое поперечное деление (архитомия); поперечное деление после дифференцировки новых систем органов (паратомия); почкование) Классификация Тип Plathelminthes (Плоские черви) кл. Turbellaria Покровы представлены ресничным эпителием Neodermata Покровы представлены погруженным синцитиальным эпителием ― тегументом (кл.Monogenoidea; кл.Trematoda; кл.Cestoda) 19. Строение турбеллярий; Класс Turbellaria (Ресничные черви) 1. Наличие ресничного эпителия. Много железистых клеток. 2. Пищеварительная система: глотка – средняя кишка (мешковидная, ветвящаяся). У представителей отр. Acoela: глотка – пищеварительная паренхима (энтодермальная). Положение ротового отверстия у разных форм сильно различается. 3. Нервная система – ортогон (у Acoela – плексус). 4. Хорошо развитые органы чувств. 5. Гермафродиты. Все кроме Acoela имеют оформленные гонады и протоки. У подотряда Neoophora (отряды Neorhabdocoela, Seriata, Tricladida и др.) имеется желточник. 20. Жизненные циклы трематод как система адаптаций; •Эндопаразиты позвоночных животных (гермафродитное поколение). •Тегумент несет шипы. У спороцист и редий тегумент формирует микроворсинки. •Марита имеет присоски: ротовая и брюшная. •Кишечник марит двуветвистый. Преобладает полостное пищеварение. •Протонефридии кроме фильтрации межклеточной жидкости способны к активному транспорту через стенки канальцев. •В состав ортогональной нервной системы входит три пары стволов. От остальных паразитических плоских червей трематоды в первую очередь отлича-ются своим сложным жизненным циклом, протекающим по типу гетерогонии. В таком цикле правильно чередуются поколения, различающиеся по способу размножения. Особи одного из них, гермафродитного, продуцируют яйцеклетки и спермии, т. е. размножаются обычным половым способом. Остальные поколения (их чаще всего два), размножаются партеногенетически; продуцируемые ими половые клетки разви-ваются без оплодотворения. Половозрелые особи гермафродитного поколения ― мариты паразитируют в позвоночных животных: рыбах, амфибиях, рептилиях, птицах и млекопитающих, в том числе и в человеке. Этот хозяин называется окончательным, или дефинитивным. Мариты по особенностям своей организации очень близки к турбелляриям, в то же время им при-сущи специфические особенности, свойственные и другим паразитическим плоским чер-вям (тегумент, устроенный по типу погруженного эпителия, наличие прикрепительного аппарата в виде присосок, и др.). Зрелые мариты продуцируют яйца, которые выводятся во внешнюю среду. Яйца по-давляющего большинства видов трематод для успешного развития должны попасть в во-ду. Под защитой яйцевой скорлупки развивается ресничная личинка первого партеногене-тического поколения ― мирацидий. Закончившие развитие мирацидии покидают яйцевые скорлупки и активно плавают в толще воды «в поисках» первого про-межуточного хозяина. Для абсолютного большинства трематод первыми промежуточными хозяевами яв-ляются моллюски ― преимущественно морские и пресноводные Gastropoda. Значительно реже в этой роли выступают наземные брюхоногие. Проникший в моллюска мирацидий претерпевает регрессивный метаморфоз (сбра-сываются ресничные эпителиальные пластинки, подвергаются дегенерации остальные личиночные органы) и превращается в материнскую спороцисту. Последняя растет, приобретает неправильную мешковидную форму тела. В ее обширной зародышевой полости в результате партеногенетического размножения формируются многочислен-ные зародыши особей следующего поколения. У разных видов трематод особи, отрождаемые материнской спороцистой, различаются по своему строению. У одних (к их числу относятся более примитивные сосальщи-ки) эти организмы называются редиями. На переднем конце их вытянутого в длину, червеобразного тела располагается ротовое отверстие, ведущее в мускулистую шаровидную глотку, за которой, в свою очередь, следует мешковидный кишечник. Основной объем тела занят обширной зародышевой полостью, в которой содержатся развивающиеся особи следующего поколения. У более специализированных трематод материнская спороциста отрождает дочерние спороцисты. Дочерние спороцисты отличаются от редий менее правиль-ной мешковидной формой тела и полным отсутствием органов пищеварительной системы. Тонкие стенки тела дочерних спороцист ограничивают обширную полость, в которой рас-полагаются многочисленные зародыши следующего поколения. И редии, и дочерние спороцисты размножаются партеногенетически ― они пред-ставляют собой второе партеногенетическое поколение в жизненном цикле сосальщиков. Редии и дочерние спороцисты могут отрождать себе подобных, т. е. снова редий и дочерних спороцист. Однако, как правило, они продуцируют свободноживущих личинок гермафродитного поколения ― церкарий. Церкарии покидают моллюска, выходят в воду и активно плавают. Для этого у них имеется специализированный локомоторный ор-ган ― хвост. Наряду с целым набором специфических личиночных органов (специальные железы, глаза и др.) церкарии обладают зачатками органов, свойственных маритам (рото-вая и брюшная присоски, зачаток пищеварительной системы, половой зачаток и т. д.). После относительно короткого периода активной жизни во внешней среде церкарии переходят в покоящуюся фазу. При этом они отбрасывают хвост и инцистируются. У бо-лее примитивных трематод личинки при этом остаются свободноживущими ― они осе-дают на субстрат, который может служить пищей для окончательного хозяина, и после этого выделяют на своей поверхности защитную оболочку ― цисту. Такие инцистиро-вавшиеся во внешней среде личинки называются адолескариями. У более специализированных сосальщиков церкарии сначала активно проникают во второго промежуточного хозяина, в роли которого могут выступать раз-личные беспозвоночные или позвоночные животные, и также инцистируются в нем. Но в этом случае их называют метацеркариями. Заражение окончательного, или дефинитивного хозяина происходит при поедании им либо адолескарий, либо инвазированных метацеркариями вторых промежуточных хозяев. В кишечнике дефинитивного хозяина личинки эксцисти-руются и в большинстве случаев остаются здесь же (кишечные виды трематод). Несколько реже они совершают миграцию к месту окончательного поселения ― в легкие, мочевой пузырь, желчные протоки и т.д. В окончательном хозяине гермафродитные особи завер-шают свой сложный метаморфоз и превращаются в марит, которые со временем достига-ют половой зрелости и приступают к откладке яиц. Таким образом, в жизненном цикле трематод, как правило, чередуются три поколе-ния: 2 партеногенетических ― материнские спороцисты и редии (или дочерние спороци-сты) и одно гермафродитное, размножающееся путем амфимиксиса. Поколение материн-ской спороцисты развивается с метаморфозом: свободноживущая личинка мирацидий, внедряясь в моллюска, превращается в материнскую спороцисту. Для редий и дочерних спороцист характерно прямое развитие без метаморфоза. Развитие особей гермафродитно-го поколения представляет собой сложный метаморфоз ― свободноживущие активно плавающие личинки церкарии превращаются в покоящихся (инцистированных) свобод-ноживущих адолескарий или паразитических метацеркарий. Адолескарии (метацеркарии) в окончательном хозяине в свою очередь превращаются в достигающую половой зрело-сти мариту. 21. Моногенеи: строение, развитие; • В основном паразиты рыб. • Как правило, эктопаразиты. •На заднем конце тела взрослых моногеней всегда имеется обособленный прикрепительный диск (присоски, крючья, многокамерные присоски, клапаны). •Пищеварительная система: глотка – двуветвистый замкнутый в кольцо кишечник. В пределах группы наблюдается постепенный переход к гематофагии. Моногенеи или моногенетические сосальщики в своем подавляющем большинстве являются паразитами рыб. Лишь некоторые представители одного таксона (отр Polyopis-thocotylidea) в качестве животных хозяев используют амфибий, рептилий и даже водных млекопитающих. Именно к этой группе и относится ставший уже классическим учебный объект Polystoma integerrimum (см. ниже). Как правило, моногенеи поселяются на поверх-ности тела своих животных-хозяев (на жабрах, плавниках и т. д.), т. е. являются экто-паразитами. Значительно реже они переходят к эндопаразитизму ― средой их оби-тания становятся, например, просвет кровеносных жаберных сосудов, полость мочевого пузыря и т. д. Будучи исходно эктопаразитами, они приобрели сложно дифференцированные и со-вершенные органы прикрепления, расположенные как на переднем, так и на заднем кон-цах тела. На переднем конце они могут быть представлены образованиями нескольких ти-пов: прикрепительными выростами, валиками, настоящими присосками. На заднем конце тела взрослых моногеней всегда имеется в той или иной степени обособленный прикрепительный диск. Последний в подавляю-щем большинстве случаев несет на своей вентральной поверхности прикрепи-тельные крючья, присоски (иногда и то, и другое одновременно) или специ-альные клапаны. В остальном моногенеи сохраняют все основные особенности орга-низации, присущие паразитическим плоским червям (Neodermata). |