Совокупность структур органов, частей органов, отдельных клеток, секретирующих в кровь и лимфу гормоны
 Скачать 1 Mb. Скачать 1 Mb.
|

|
В средней части аденогипофиза вырабатывается меланоцитостимулирующий гормон (меланоцитотропин), а также липотропин — гормон, усиливающий метаболизм липидов. Туберальная часть аденогипофиза — отдел, прилежащий к гипофизарной ножке и соприкасающийся с нижней поверхностью медиального возвышения гипоталамуса. 13. Охарактеризуйте особенности кровоснабжения гипофиза. Гипоталамо-аденогипофизарное кровоснабжение и его значение в гипоталамической регуляции аденогипофизарного гормонопоэза (см. рис. 15.2). Система гипоталамо-аденогипофизарного кровоснабжения называется портальной (воротной). Приносящие гипофизарные артерии вступают в медиальное возвышение медиобазального гипоталамуса, где разветвляются в сеть капилляров (первичное капиллярное Сплетение портальной системы). Эти капилляры образуют петли и клубочки, с которыми контактируют терминали аксонов нейросекреторных клеток аденогипофизотропной зоны гипоталамуса. Капилляры первичного сплетения собираются в портальные вены, идущие вдоль гипофизарной ножки в переднюю долю, где они распадаются на капилляры синусоидного типа (вторичная капиллярная сеть), разветвляющиеся между трабекулами паренхимы железы. Наконец, синусоиды вторичной капиллярной сети собираются в выносящие вены, по которым кровь, обогатившаяся гормонами передней доли, поступает в общую циркуляцию. 14. Охарактеризуйте нейрогемальные контактные области и приведите их примеры. Нейроэндокринные трансдукторы (переключатели) и нейрогемальные образования. Медиальное возвышение (eminentia medialis) является нейрогемальным органом гипоталамо-аденогипофизарной системы. Оно образовано эпендимой, отдельные глиальные клетки которой дифференцируются в танициты (tanicyti), отличающиеся разветвленными отростками, контактирующими с клубочками первичной капиллярной сети портальной системы гипоталамогипофизарного кровообращения. В гипоталамо-аденогипофизарной системе аккумулируются аденогипофизотропные нейрогормоны — нейротрансмиттеры (либерины и статины), вырабатываемые в мелкоклеточных ядрах среднего и заднего отделов гипоталамуса, которые затем поступают в портальную систему гипофиза. В гипоталамо-нейрогипофизарной системе аналогичным нейрогемальным органом оказывается нейрогипофиз (задняя доля гипофиза), где аккумулируются нонапептидные нейрогормоны (вазопрессин — антидиуретический гормон и окситоцин), вырабатываемые в крупноклеточных ядрах переднего отдела гипоталамуса, в дальнейшем выделяемые в кровь. 15. Опишите компоненты и дайте функциональную характеристику гипоталамо-гипофизарных связей. Аксоны мелких пептидоадренергических клеток, равно как и сопровождающих их адренергических нейронов обычного типа, направляются в медиальную эминенцию в составе тубероиНфундибулярного дофаминергического пучка, берущего начало от дофаминергических нейронов туберальных ядер (рис. 15.2). 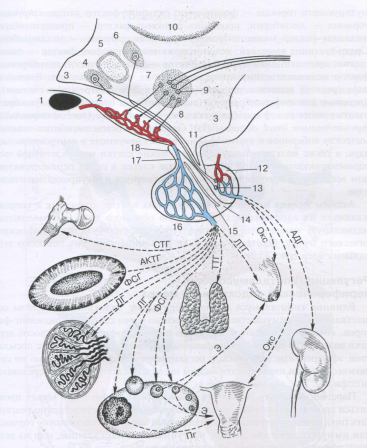   16. Перечислите железы, являющиеся производными глоточных карманов. ПЕРИФЕРИЧЕСКИЕ ЭНДОКРИННЫЕ ЖЕЛЕЗЫ Бранхиогенная группа эндокринных желез развивается из зачатков жаберных карманов и включает щитовидную и околощитовидные железы. Из зачатков жаберных карманов развивается также тимус. Щитовидная железа и околощитовидные железы связаны не только общим источником развития, но и функционально, играя главную роль в поддержании метаболического статуса и постоянства состава внутренней среды организма. Гормоны этих желез (тироксин, кальцитонин, паратгормон) регулируют скорость основного обмена и концентрацию кальция в крови. 17. Опишите общую структуру щитовидной железы и назовите ее структурно-функциональную единицу. Щитовидная железа (glandula thyroidea) содержит два клеточных дифферона эндокриноцитов, развивающихся из стволовых клеток различного происхождения: Т-тироциты (фолликулярные клетки), наиболее многочисленные, вырабатывающие йодсодержащие гормоны — тироксин (Т4) и трийодтиронин (Т3), и С-тироциты (парафолликулярные клетки), вырабатывающие гормон тирокальцитонин и ряд других пептидов. Развитие. Зачаток щитовидной железы возникает у зародыша человека на 3—4-й нед как выпячивание стенки глотки между I и II парами жаберных карманов, которое растет вдоль глоточной киш ки в виде эпителиального тяжа. На уровне III—IV пары жаберных карманов этот тяж раздваивается, давая начало формирующимся правой и левой долям щитовидной железы. Начальный эпителиальный тяж (ductus thyreoglossus) атрофируется, и от него сохраняются только перешеек, связывающий обе доли щитовидной железы, и проксимальная часть в виде ямки {foramen соесит) в корне языка. У большинства других млекопитающих атрофируется также дистальный конец эпителиального тяжа, поэтому перешеек не развивается и обе доли щитовидной железы обособляются. Зачатки долей быстро разрастаются, образуя рыхлые сети ветвящихся эпителиальных трабекул, клетки которых дифференцируются в Т-тироциты. Последние объединяются в фолликулы, в промежутки между которыми врастает мезенхима с кровеносными сосудами и нервами. По мере развития в зачаток железы врастают также производные V пары жаберных карманов — клетки так называемых ультимобранхиальных телец. Это С-тироциты, которые имеют нейроэктодермальную природу, и в закладку щитовидной железы они внедряются через ультимобранхиальные зачатки. Строение. Щитовидная железа окружена соединительнотканной капсулой, прослойки которой направляются вглубь и разделяют орган на дольки, в которых располагаются многочисленные сосуды микроциркуляторного русла и нервы. Основными структурными компонентами паренхимы железы являются фолликулы. 18. Назовите эпителиальные клеточные элементы щитовидной железы и их функции. Фолликулы — замкнутые шаровидные или слегка вытянутые образования варьирующих размеров с полостью внутри, выстланные одним слоем эпителиальных клеток, представленных Т-тироцитами (фолликулярными клетками), а также С-тироцитами (парафолликулярными клетками) нейрального происхождения (рис. 15.8). В дольке железы различают фолликулярные комплексы (микродольки), которые состоят из группы фолликулов, окруженных тонкой соединительнотканной капсулой. В просвете фолликулов накапливается коллоид — секреторный продукт Т-тироцитов, представляющий собой вязкую жидкость, состоящую в основном из тироглобулина. Размер фолликулов и образующих их стенку Т-тироцитов варьирует в нормальных физиологических условиях. Их диаметр от 0,02 до 0,9 мм. В небольших формирующихся фолликулах, еще не заполненных коллоидом, эпителий однослойный призматический. По мере накопления коллоида размеры фолликулов увеличиваются, эпителий становится кубическим, а в сильно растянутых фолликулах, заполненных коллоидом, — плоским. Основная масса фолликулов в норме выстлана Т-тироцитами кубической формы. Увеличение размеров фолликулов обусловлено пролиферацией, ростом и дифференцировкой Т-тироцитов, сопровождаемой накоплением коллоида в полости фолликула. Фолликулы разделяются тонкими прослойками рыхлой волокнистой соединительной ткани с многочисленными кровеносными и лимфатическими капиллярами, оплетающими фолликулы, тучными клетками и лимфоцитами (рис. 15.9). Т-тироциты, или фолликулярные эндокриноциты (endocrinocytus follicular is), — железистые клетки, составляющие большую часть стенки фолликулов. В фолликулах Т-тироциты образуют выстилку (стенку) и располагаются в один слой на базальной мембране. При умеренной функциональной активности щитовидной железы (ее нормальной функции) Т-тироциты имеют кубическую форму и шаровидные ядра. Коллоид, секретируемый ими, заполняет в виде гомогенной массы просвет фолликула. На апикальной поверхности Т-тироцитов, обращенной к просвету фолликула, имеются микроворсинки. По мере усиления тиреоидной активности количество и размеры микроворсинок возрастают. Одновременно базальная поверхность Т-тироцитов, почти гладкая в период функционального покоя щитовидной железы, становится складчатой, что увеличивает соприкосновение Т-тироцитов с перифолликулярными пространствами. Соседние клетки в выстилке фолликулов тесно связаны между собой многочисленными десмосомами и хорошо развитыми терминальными пластинками. По мере возрастания тиреоидной активности на боковых поверхностях Т-тироцитов возникают пальцевидные выступы (интердигитации), входящие в соответствующие вдавления боковой поверхности соседних клеток. 19. Опишите основные структурные изменения щитовидной железы при гипо- и гиперфункции. В Т-тироцитах хорошо развиты органеллы, особенно участвующие в белковом синтезе. Белковые продукты, синтезируемые Т-тироцитами, выделяются в полость фолликула, где завершается образование йодированных тирозинов и тиронинов (аминокислот, входящих в состав крупной и сложной молекулы тироглобулина). Тиреоидные гормоны могут попасть в циркуляцию лишь после высвобождения из этой молекулы (т. е. после расщепления тироглобулина). Когда потребности организма в тиреоидном гормоне возрастают и функциональная активность щитовидной железы усиливается (гиперфункция щитовидной железы), Т-тироциты фолликулов принимают призматическую форму. Интрафолликулярный коллоид при этом становится более жидким и содержит многочисленные ресорбционные вакуоли. Ослабление функциональной активности (гипофункция щитовидной железы) проявляется, наоборот, уплотнением коллоида, его застоем Внутри фолликулов, диаметр и объем которых значительно увеличиваются; высота же Т-тироцитов уменьшается, они принимают уплощенную форму, а их ядра вытягиваются параллельно поверхности фолликула. 20. Опишите секреторный цикл щитовидной железы. Секреторный цикл. В секреторном цикле различают основные фазы: фазу продукции и фазу выведения гормонов (рис. 15.10). Фаза продукции включает: 1) поступление предшественников тироглобулина (аминокислот, углеводов, ионов, воды, йодидов), приносимых из кровеносного русла в Т-тироциты; 2) синтез полипептидных цепочек тироглобулина в гранулярной эндоплазматической сети и их гликозилирование (соединение с нейтральными сахарами и сиаловой кислотой) с помощью фермента тиропероксидазы в комплексе Гольджи; синтез тиропероксидазы, окисляющей йодиды и обеспечивающей их соединение с тироглобулином на поверхности Т-тироцитов и образование коллоида (йодирование тироглобулина). При этом к нейодированному тироглобулину присоединяется сначала один атом йода, а затем и второй, в результате чего образуются моно- и дийодтиронины. Последующая их комплексация дает трийодтиронин и тетрайодтиронин (тироксин). Фаза выведения включает эндоцитоз тиреоидных гормонов, связанных с тироглобулином, которые подвергаются гидролизу с помощью лизосомных протеаз. Тироглобулин расщепляется до аминокислот, а монойодтирозин, дийодтирозин, трийодтиронин (Т3) и тетрайодтиронин (Т4) освобождаются в цитоплазму. Два последних выводятся через базальную мембрану в капилляры и лимфатические капилляры, а монойодтирозин и дийодтирозин используются для синтеза новой молекулы тироглобулина. Тиротропный гормон усиливает функцию щитовидной железы, стимулируя поглощение тироглобулина микроворсинками Т-тироцитов, а также его расщепление в фаголизосомах с высвобождением активных гормонов. С-тироциты, или парафолликулярные эндокриноциты (endocrinocytus parafollicularis). Во взрослом организме парафолликулярные эндокринные клетки локализуются в стенке фолликулов, залегая между основаниями соседних Т-тироцитов, но не достигают своей верхушкой просвета фолликула (интраэпителиальная локализация парафолликулярных клеток). Кроме того, парафолликулярные клетки располагаются также в межфолликулярных прослойках соединительной ткани (рис. 15.11). По размерам парафолликулярные клетки крупнее Т-тироцитов, имеют округлую, иногда угловатую форму. В отличие от Т-тироцитов парафолликулярные клетки не поглощают йод, но совмещают образование нейроаминов (норадреналина и серотонина) путем декарбоксилирования тирозина и 5-гидрокситриптофана (ароматических аминокислот — предшественников указанных нейроаминов) с биосинтезом белковых (олигопептидных) гормонов — кальцитонина и соматостатина. Секреторные гранулы, густо заполняющие цитоплазму парафолликулярных клеток, обнаруживают сильную осмиофилию и аргирофилию. В цитоплазме парафолликулярных клеток хорошо развиты гранулярная эндоплазматическая сеть и комплекс Гольджи. Секреторные гранулы парафолликулярных клеток бывают двух типов. В некоторых парафолликулярных клетках преобладают мелкие, но сильно осмиофильные гранулы. Клетки данного типа вырабатывают кальцитонин. В парафолликулярных клетках другого типа содержатся более крупные, но слабо осмиофильные гранулы. Эти клетки продуцируют соматостатин (рис. 15.12). 21. Опишите общую структуру, клеточный состав и функции околощитовидных желёз. Околощитовидные железы (glandulaeparathyroideae) (4—5) расположены на задней поверхности щитовидной железы и отделены от нее капсулой. Масса желез 0,05—0,3 г. Функциональное значение околощитовидных желез заключается в регуляции метаболизма кальция. Они вырабатывают белковый гормон паратирин, который стимулирует резорбцию кости остеокластами, повышая содержание кальция в крови, и снижает содержание фосфора в крови, тормозя его резорбцию в почках, уменьшает экскрецию кальция почками, усиливает синтез 1—2,5-дигидроксихолекальциферола (метаболита витамина D), который повышает содержание кальция в сыворотке и его всасывание в пищеварительном тракте. Паратирин и кальцитонин тесно взаимодействуют в регуляции минерального обмена: кальцитонин снижает уровень кальция в крови; паратирин является антагонистом кальцйтонина. Гипокальциемия усиливает секрецию паратирина, а гиперкальциемия, наоборот, подавляет. Кальцитонин и паратирин также действуют на функцию почек и пищеварительный тракт, регулируя экскрецию и поглощение кальция в этих органах. Развитие. Околощитовидные железы закладываются у зародыша как выступы из эпителия III и IV пары жаберных карманов глоточной кишки. На 5—6-й нед эмбриогенеза образуются четыре зачатка желез в виде эпителиальных почек. На 7—8-й нед эти почки отшнуровываются от стенок жаберных карманов, присоединяясь к задней поверхности щитовидной железы. В процессе гистогенеза эпителия околощитовидных желез составляющие его клетки становятся все более дифференцированными, их размеры увеличиваются, количество гликогена в них уменьшается, цитоплазма приобретает светлую окраску. Они называются главными паратироцитами. У 5-месячного плода главные паратироциты дифференцируются на светлые и темные паратироциты. На 10-м году жизни появляется следующий вид эпителиальных клеток желез — ацидофильные, или оксифильные, паратироциты. В виде единичных включений в паренхиме околощитовидных желез могут находиться С-клетки, вырабатывающие кальцитонин. Строение. Каждая околощитовидная железа окружена тонкой соединительнотканной капсулой. Ее паренхима представлена трабекулами — эпителиальными тяжами либо скоплениями эпителиальных эндокринных клеток — паратироцитов (parathyrocytus endocrinus), разделенными тонкими прослойками рыхлой соединительной ткани с многочисленными капиллярами (рис. 15.13, а, б). Хотя между паратироцитами хорошо развиты межклеточные щели, соседние клетки связаны интердигитациями и десмосомами. Различают главные паратироциты (parathyrocytus principalis) и оксифильные паратироциты (parathyrocytus oxyphilicus). Главные клетки секретируют паратирин, они преобладают в паренхиме железы, имеют небольшие размеры (7—10 мкм), полигональную форму. В периферических зонах цитоплазма базофильна, где рассеяны скопления свободных рибосом (полисом). Секреторные гранулы имеют диаметр 150—200 нм. При усилении секреторной активности околощитовидных желез главные клетки увеличиваются в объеме. Среди главных паратироцитов различают светлые (parathyrocytus principalis lucidus) неактивные формы и темные (parathyrocytus principalis densus) активные формы. В последних хорошо развиты органеллы синтеза секрета. В цитоплазме светлых клеток встречаются включения гликогена и жира. Обычно на один темный приходится 3—5 светлых паратироцитов. Оксифилъные паратироциты малочисленны, располагаются поодиночке или группами, они значительно крупнее, чем главные паратироциты. В цитоплазме видны оксифильные гранулы, огромное количество митохондрий, слабо развитый комплекс Гольджи. Их рассматривают как стареющие формы главных клеток. Другая точка зрения состоит в том, что это клетки APUD-серии. Выделяют также промежуточный тип клеток. На секреторную активность околощитовйдных желез не оказывают влияния гипофизарные гормоны. Околощитовидная железа по принципу обратной связи быстро реагирует на малейшие колебания содержания кальция в крови. Ее деятельность усиливается при гипокальциемии и ослабляется при гиперкальциемии. Паратироциты обладают рецепторами, способными непосредственно воспринимать прямые влияния на них ионов кальция. 22. Опишите источник развития, структуру и функциональные характеристики эпифиза. Эпифиз — верхний мозговой придаток (epiphysis cerebri), или шишковидное тело (corpus pineale). Эпифиз участвует в регуляции процессов, протекающих в организме ритмически или циклически, например овариально-менструального цикла. Ритмические колебания других периодических функций, интенсивность которых закономерно изменяется на протяжении суток, называются циркадными (от лат. circa diem — около дня). Циркадные ритмы явно связаны со сменой дня и ночи (светового и темнового периодов), и их зависимость от эпифиза свидетельствует о том, что гормонообразовательная деятельность последнего определяется его способностью различать смену световых раздражений, получаемых организмом. Развитие. У зародыша человека эпифиз развивается как выпячивание крыши III желудочка промежуточного мозга на 5—6-й нед развития. В его состав включается субкомиссуральный орган, который развивается из эпендимы III желудочка мозга. У человека и млекопитающих он сильно редуцирован (0,2 г). В результате дивергентной дифференцировки нейральных стволовых клеток развиваются два клеточных дифферона — пинеалоцитарный и глиоцитарный. Максимального развития эпифиз достигает у детей до 7 лет. |