лол. Спинной мозг. Структурнофункциональные особенности серого вещества. Сегментарный тип строения, двигательные центры. Характеристика спинальных нейронов, их функциональное значение.
 Скачать 110.98 Kb. Скачать 110.98 Kb.
|

|
Спинной мозг. Структурно-функциональные особенности серого вещества. Сегментарный тип строения, двигательные центры. Характеристика спинальных нейронов, их функциональное значение. Возвратное торможение в спинном мозге. Эфферентные нейроны вегетативной нервной системы. 1) Спинной мозг --- часть ЦНС, заключенная в позвоночный столб. Это самая древняя часть ЦНС, самый низший ее отдел. Имеет сегментарное строение, состоит из 31 сегмента. Сегмент – это участок спинного мозга, который включает две пары корешков и участвует в иннервации трех метамеров тела. Имеет 2 утолщения: шейное и поясничное. Нейроны спинного мозга образуют его серое вещество в виде симметрично расположенных 2 передних и 2 задних рогов. В сером веществе нейроны образуют ядра – скопления однородных по функциям нейронов. Задние корешки являются афферентными, чувствительными, центростремительными, а передние — эфферентными, двигательными, центробежными (закон Белла—Мажанди). Задние рога выполняют главным образом сенсорные функции и содержат интернейроны, функция которых заключается в организации связей между структурами спинного мозга, в обеспечении влияния восходящих и нисходящих путей на клетки отдельных сегментов спинного мозга, а также торможение активности нейронов с сохранением направленности пути возбуждения. Передние рога выполняют двигательную функцию, в них располагаются альфа- и гамма-мотонейроны. Альфа-мотонейроны иннервируют экстрафузальные мышечные волокна, а гамма-мотонейроны – интрафузальные мышечные волокна, которые содержат рецепторы растяжения. Двигательные центры --- совокупность нейронов ЦНС: - двигательные центры (иннервируют по сегментарному принципу скел мускулатуру) - центры симпатического и парасимпатического отдела В тораколюмбальном отделе (C8-L2) имеется боковые рога, содержащие симпатические нейроны, в S2-S4 парасимпатические. Все, что расположено выше сигмовидной кишки иннервируется парасимпатической частью ЦНС, все, что ниже – симпатической. Функционально нейроны спинного мозга можно разделить на 4 основные группы: По принадлежности к отделу н/с - соматические: - вегетативные: симпатические и парасимпатические нейроны расположены преимущественно в боковых рогах; аксоны этих нейронов выходят из спинного мозга в составе передних корешков; 2) По назначению: - Эфферентные - мотонейроны, или двигательные, — клетки передних рогов, аксоны которых образуют передние корешки; альфа1 – быстрые, иннервирующие белые мыш вол альфа1 – медленные, иннервирующие красные мыш вол гамма – иннервируют интрафузальные мышечные волокна, которые содержат рецепторы растяжения - Афферентные --- локализуются в спинальных ганглиях и ганглиях чер нервов. Их отростки, проводящие афферентную импульсацию от мыш-х, сух-х и кожных рецепторов, вступают в ствол мозга или соответствующие сегменты спин мозга и образуют синаптические контакты либо на альфа-мотонейронах, либо на вставочных. - Интернейроны — нейроны, получающие информацию от спинальных ганглиев и располагающиеся в задних рогах; реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения. Устанавливают связь с мотонейронами спин мозга аф-х нейронов, с ядрами ствола мозга. Могут быть возбуждающими и тормозными (клетки Реншоу) - Ассоциативные клетки — нейроны собственного аппарата спинного мозга, устанавливающие связи внутри и между сегментами, поддержание позы, тонуса мышц, движений. 3) Возвратным торможением называется угнетение (подавление) активности нейрона, вызываемое возвратной коллатералью аксона нервной клетки. Так, мотонейрон переднего рога спинного мозга прежде чем покинуть спинной мозг дает боковую (возвратную) ветвь, которая возвращается назад и заканчивается на тормозных нейронах (клетки Реншоу). Аксон последней заканчивается на мотонейронах, оказывая на них тормозное действие. 4) Эфферентные нейроны нервной системы — это нейроны, передающие информацию от нервного центра к исполнительным органам или другим центрам нервной системы. α -мотонейрон иннервирует экстрафузальные мышечные волокна – медленные двигательные единицы (обыч мышцы) и характеризуются низкой частотой импульсации. Обеспечивают тоническое сокращение скел мышц. Они получают информацию от чувствительных нейронов. γ –мотонейроны иннервируют интрафузальные МВ, которые содержат рецепторы растяжения или мышечные веретена. Они тоньше и короче и располагаются параллельно экстрафузальным. Вокруг средней части имеются афферентные волокна (анулоспинальные окончания)- толстые миелинизированные волокна А- α типа, они разветвляются и образуют первичные окончания мышечных веретен. Интрафузальные МВ, кроме афферентной иннервации имеют эфферентную иннервацию, которую они получают от γ-мотонейронов. γ -мотонейроны имеют более тонкие миелинизировные аксоны, чем α- мотонейроны. Чем более тонкие движения совершает МВ, тем больше плотность мышечных веретен на единицу мыш поверхности. Рефлекторная деятельность спинного мозга. Сухожильные и кожные рефлексы, их значение. Понятие о гамма-петле.. Двигательные рефлексы спинного мозга (сгибательные, разгибательные, локомоторные, перекрестно-разгибательные), механизм их возникновения и физиологическое значение. 1) Собственная рефлекторная деятельность СМ осуществляется сегментарными рефлекторными дугами. Сегментарная рефлекторная дуга состоит из рецептивного поля, от которого импульсация по чувствительному волокну спинального ганглия, а затем по аксону этого же нейрона через задний корешок входит в спинной мозг, далее аксон может идти прямо е мотонейрону переднего рога аксон которого подходит к мышце. Это моносинаптическая рефлекторная дуга. С участием вставочных нейронов – полисинаптическая. Рефлексы: •Сухожильные рефлексы - это рефлексы, возникающие при механическом раздражении мышц и сухожилий, небольшое растяжение которых приводит к возбуждению рецепторов растяжения, после чего сигналы от альфа-мотонейронов спинного мозга поступают к мышцам и способствуют их сокращению. К ним относятся коленный, ахиллов, локтевой, кистевой и др. рефлексы. К интрафузальным мышечным волокнам (интерорецепторам) относятся мышечные веретёна и сухожильные органы Гольджи. Мышечные веретна располагаются параллельно экстрафузальным (скелетным) мышечным волокнам, прикрепляясь одним концом к скелетному мышечному волокну, а другим – к сухожилию мышцы; Функция: Мышечные веретёна являются рецепторами растяжения; они предохраняют мышцу от перерастяжения, обеспечивая периферический тонус мышц (слабо возбуждаются при расслаблении мышцы и сильно при её растяжении). Механизм работы мышечных веретён: Скелетная мышца расслабляется, вследствие чего возбуждается мышечное веретено возбуждаются первичные афференты (Ia-волокна), по которым сигнал передаётся в спинной мозг (Ia-волокна образуют синапсы с альфа-мотонейронами, иннервирующими ту мышцу, от мышечного веретена которой поступил сигнал) альфа-мотонейрон возбуждается, способствуя сокращению мышцы, вследствие чего её растяжение уменьшается. Сухожильный орган Гольджи соединяются со скелетным мышечным волокном ПОСЛЕДОВАТЕЛЬНО, возбуждаясь при сокращении мышцы; Функция: Сухожильные органы Гольджи являются датчиками напряжения; они служат для поддержания постоянства напряжения мышцы. Механизм работы сухожильных органов Гольджи: При сокращении экстрафузальных (скелетных) мышечных волокон, увеличивается напряжение мышцы, что приводит к возбуждению афферентов Ib эти афференты идут в спинной мозг, где образуют синапсы со вставочными нейронами – клетками Реншоу (тормозные нейроны спинного мозга) клетка Реншоу образует тормозной синапс на альфа-мотонейронах, иннервирующих ту же скелетную мышцу, от сухожильного органа которой пришёл сигнал. При этом происходит подавление активности данного мотонейрона и мышца расслабляется. Возможен и противоположный механизм: при ослаблении мышечного тонуса мотонейроны РАСТОРМАЖИВАЮТСЯ (уменьшается степень их торможения) и тем самым активируются. •Кожные рефлексы--- это рефлексы, обусловленные раздражением рецепторов кожи, но проявляющиеся двигательными реакциями. Ими являются подошвенный, брюшной и кремастерный рефлексы Механизм центрального тонуса мышцы (гамма-петля): Сигнал с ретикулярной формации передаётся на гамма-мотонейрон, иннервирующий мышечное веретено происходит сокращение мышечных волокон мышечного веретена возбуждаются Ia-волокна мышечного веретена, по которым сигнал передаётся в спинной мозг на альфа мотонейроны, иннервирующие ту мышцу, от мышечного веретена которой пришёл сигнал по гамма-мотонейрону происходит сокращение мышцы. 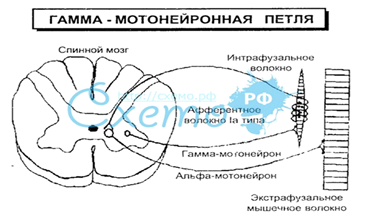 3) Двигательные рефлексы спинного мозга (сгибательные, разгибательные, локомоторные, перекрестно-разгибательные), механизм их возникновения и физиологическое значение. • Сгибательные рефлексы --- однократное сгибание конечности при однократном раздражении кожи или проприорецепторов. Сгибательные рефлексы направлены на избежание различных повреждающих воздействий и возникают при раздражении болевых рецепторов кожи, мышц и внутренних органов. Если вызвать болевое раздражение конечности спинального животного, она отдёргивается путём сгибания в суставах данной конечности, что обеспечит отдаление конечности от источника болевого стимула. Из этого следует, что сгибательный рефлекс является ОБОРОНИТЕЛЬНЫМ РЕФЛЕКСОМ. • Разгибательные рефлексы возникают только с проприорецепторов мышц-разгибателей и являются моносинаптическими. Они (как и сгибательные) подразделяются на фазные (сокращение мышцы при однократном раздражении кожи или проприорецепторов) и тонические (направленные на поддержание позы и осуществляемые с помощью проприорецепторов). Приверы: коленный • Локомоторные рефлексы. Локомоция – это перемещение человека или животного в пространстве при помощи координированных движений конечностей. Она запрограммирована на уровне спинного мозга. Болевое раздражение какой-либо конечности спинального животного вызывает рефлекторные движения всех четырёх конечностей. Если такая стимуляция продолжается долго, могут возникнуть ритмичные сгибательные и разгибательные движения не подвергающихся раздражению конечностей. • Перекрёстно-разгибательные рефлексы – это рефлексы, при которых сгибание одной конечности сопровождается разгибанием противоположной, особенно при болевом раздражении. При этом импульсы от болевых волокон переходят на противоположную сторону спинного мозга, запуская там реакцию разгибания. Проводниковая деятельность спинного мозга. Характеристика афферентной импульсации, поступающей по восходящим путям к структурам головного мозга. Нисходящие проводящие пути, их основные физиологические функции. Последствия поперечной травмы спинного мозга на разных уровнях. Явление спинального шока. 1) Проводниковая функция спинного мозга. Аксоны спинальных ганглиев и нейронов серого вещества спинного мозга идут в его белое вещество и в другие структуры ЦНС, создавая проводящие пути. Восходящие пути спинного мозга: • Пучки Голля (тонкий) и Бурдаха (клиновидный) – проприоцептивные пути, начинающиеся от рецепторов глубокой чувствительности мышц, сухожилий, надкостницы, оболочек суставов. Тонкий пучок (Голля) начинается от ганглиев, собирающих информацию от нижней половины тела (19 нижних сегментов спинного мозга). Клиновидный пучок (Бурдаха) начинается от ганглиев, собирающих информацию от верхней части тела (12 верхних сегментов спинного мозга). • Спинноталамический путь - основной путь кожной чувствительности. Он начинается от болевых, температурных, тактильных рецепторов и барорецепторов кожи, сигналы от которых идут к нейронам спинального ганглия, а далее через задний корешок к заднему рогу спинного мозга. Здесь происходит ПЕРВОЕ переключение на новый нейрон, расположенный в собственных ядрах заднего рога. Отсюда путь идёт на ПРОТИВОПОЛОЖНУЮ сторону спинного мозга и по боковому канатику поднимается к таламусу (коллектору всех видов общей чувствительности). От таламуса путь направляется в сенсорную область коры большого мозга (в постцентральную извилину). Скорость проведения возбуждения по волокнам данного тракта – 20-30 м/с. • Спинномозжечковые пути лежат в боковых канатиках спинного мозга и представлены НЕПЕРЕКРЕЩИВАЮЩИМИСЯ передним спинно-мозжечковым путём (пучок Говерса) и ДВАЖДЫ ПЕРЕКРЕЩИВАЮЩИМСЯ задним спинно-мозжечковым путём (пучок Флексига). Следовательно, мозжечок получает информацию, которая идёт от сухожильных рецепторов Гольджи, проприорецепторов, рецепторов давления, прикосновения, в обоих случаях от своей половины тела. Эти пути проводят импульсы бессознательной проприоцептивной чувствительности. Скорость проведения возбуждения по волокнам данного тракта – 110-120 м/с. Нисходящие пути: • Пирамидный путь состоит из двух пучков: латерального и прямого, начинающихся от клеток Беца двигательной зоны коры больших полушарий головного мозга. Латеральный пучок на уровне продолговатого мозга переходит на ПРОТИВОПОЛОЖНУЮ сторону и спускается к мотонейронам определённого сегмента спинного мозга. Прямой пучок спускается до своего сегмента и там переходит к мотонейронам ПРОТИВОПОЛОЖНОЙ стороны. Следовательно, перекрещивающимся является весь пирамидный путь. Его функция заключается в проведении импульсов сознательного управления скелетной мускулатурой. Скорость проведения возбуждения по волокнам данного тракта – 50-100 м/с. • Руброспинальный путь состоит из аксонов нейронов красного ядра, которые по выходе из данной структуры головного мозга сразу образуют ПЕРЕКРЁСТ и делятся на 3 пучка: к спинному мозгу, в мозжечок, в ретикулярную формацию ствола мозга. Нейроны, идущие в спинной мозг участвуют в управлении мышечным тонусом, а остальные обеспечивают координацию активности пирамидных нейронов коры и нейронов мозжечка, участвующих в организации произвольных движений. Скорость проведения возбуждения по волокнам данного тракта – 60-80 м/с. • Вестибулоспинальный путь начинается от нейронов латерального преддверного ядра (ядра Дейтерса), лежащего в продолговатом мозге, идёт в спинной мозг, заканчиваясь посегментно на двигательных анимальных ядрах передних рогов спинного мозга. Он регулирует активность мотонейронов спинного мозга, обеспечивает тонус мускулатуры, согласованность движения, равновесие. Он осуществляет рефлекторные двигательные реакции в ответ на неожиданные слуховые раздражения (страж-рефлекс по Павлову). Скорость проведения возбуждения по волокнам данного тракта – 50-60 м/с. • Ретикулоспинальный тракт идёт от нейронов ретикулярной формации ствола мозга к мотонейронам спинного мозга. Через него ретикулярная формация регулирует тонус мускулатуры. Скорость проведения возбуждения по волокнам данного тракта – 30-40 м/с. 2) При поперечной травме спинного мозга: • выше 4 шейного позвонка – смерть, т.к. повреждаются диафрагмальные нервы; • на уровне грудного отдела – паралич всех конечностей; • на уровне поясничного отдела – паралич нижних конечностей. После полного пересечения или травматического разрыва спинного мозга возникает спинальный шок, проявляющийся в отсутствии чувствительности, параличе всех произвольных движений мышц и угнетении двигательных и вегетативных рефлексов ниже места повреждения. Спинальный шок (по лекции) --- отсутствие сухожильных рефлексов, переходящее в угнетение рефлекторной активности спинного мозга ниже уровня травмы с утратой двигательных, чувствительных и вегетативных функций. Продолжительность от нескольких недель до нескольких месяцев. Рефлексы могут угнетаться обратимо (дисрефлексия) или полностью отсуствуют (арефлексия). Причиной арефлексии является то, что нейроны СМ, в норме постоянно получающие сигналы из ГМ, после повреждения оказываются гиперполяризованными. Восстановление происходит медленно. В последнюю очередь восстанавливаются моносинаптические рефлекторные дуги. Произвольные движения после полного пересечения СМ практически не восстанавливаются. ???????????????? Продолговатый мозг и варолиев мост. Жизненно-важные центры продолговатого мозга. Рефлексы продолговатого мозга (двигательные, висцеральные, позно-тонические, вестибулярные, шейные), их характеристика. Статические (рефлексы положения, выпрямления) и статокинетические рефлексы, механизм образования, их значение. Проводниковая функция продолговатого мозга. Участие варолиева моста в механизме сна. У человека достигает 25 мм в длину. Является продолжением СМ. Не имеет метамерного принципа строения, серое вещество в нем расположено не в центре, а в виде ядер. В ПМ имеются оливы, связанные со СМ, экстрапирамидной системой и мозжечком, ядра Голля и Бурдаха, а также ретикулярная формация. Продолговатый мозг регулирует ряд сенсорных функций с помощью расположенных в нём ядер: ядро языкоглоточного нерва обеспечивает первичный анализ рецепции вкуса; ядро улиткового нерва обеспечивает рецепцию слуховых раздражений; верхнее вестибулярное ядро обеспечивает рецепции вестибулярных раздражений; ядро блуждающего нерва обеспечивает восприятие чувствительной информации от многих внутренних органов, а также отдаёт к ним вегетативные волокна. Кроме того, двигательное ядро обеспечивает последовательное сокращение мышц глотки, гортани при глотании; тонкое и клиновидное ядра обеспечивает переключение на второй нейрон волокон путей Голля и Бурдаха. Ядро подъязычного (12 пара), добавочного (11 пара) - двигательные. Ядра блуждающего и языкоглоточного нерва. 2) Жизненно важные центры, расположенные в ПМ: 1) Дыхательный центр. Имеет 2 отдела: инспираторный, отвечающий за вдох и экспираторный, отвечающий за выдох. Афферентные сигналы в дыхательный центр поступают от диафрагмы, межреберных мышц, верхних дыхательных путей, рецепторов альвеол и сосудов. Эфферентные волокна идут к мотонейронам, которые иннервируют диафрагму и межреберные мышцы. Нейроны дыхательного центра способны реагировать на уровень кислорода, углекислого газа рН крови. Характерная особенность дыхательного центра – автоматия, т.е нейроны дыхательного центра способны к самовозбуждению. 2) Сосудодвигательный центр. Афферентные сигналы поступают от рецепторов сосудов, бронхиол сердца, органов брюшной полости. Эфферентные сигналы – по ретикулоспинальному тракту к боковым рогам СМ. Он функционирует совместно с вышележащими структурами мозга, и прежде всего с гипоталамусом. Возбуждение сосудодвигательного центра всегда изменяет ритм дыхания, тонус бронхов, мышц кишечника, мочевого пузыря, цилиарной мышцы и др. Это обусловлено тем, что ретикулярная формация продолговатого мозга имеет синаптические связи с гипоталамусом и другими центрами. 3) Центр жевания 4) Центр сосания 5) Центр глотания 6) Центр чихания 7) Центр кашля 8) Центр слюноотделения 9) Центр мигания 10) Главное ядро пищеварительной системы, которое отвечает за секреторную и моторную деятельность пищ-го тракта. 11) Центр защитных рефлексов: кашель, рвота, чихание, слезы. 12) Центры сложных двигательных рефлексов 13) Ретикулярная фармация, которая активирует дыхание и сосудодвигательный центр. 5) Проводниковые функции. Через продолговатый мозг проходят все восходящие и нисходящие пути спинного мозга: спино-таламический, кортикоспинальный, руброспинальный. В нем берут начало вестибулоспинальный, оливоспинальный и ретикулоспинальный тракты, обеспечивающие тонус и координацию мышечных реакций. В продолговатом мозге заканчиваются пути из коры большого мозга — корково-ретикулярные пути. Здесь заканчиваются восходящие пути проприоцептивной чувствительности из спинного мозга: тонкого и клиновидного. Рефлексы продолговатого мозга (двигательные, висцеральные, позно-тонические, вестибулярные, шейные), их характеристика Тонические – рефлексы, которые направлены на сохранение естественной позы, на определенную ориентацию тела в пространстве. Возникают в покое и могут возникать при перемещении тела в пространстве. Рефлексы позы: 1) статические: Рефлексы положения. Это изменение тонуса мышц при перемене положения тела в пространстве. Рефлексы выпрямления. Перераспределение тонуса мышц, приводящих к восстановлению естественной позы в случае ее изменения. В осуществлении этих рефлексов принимают участие вестибулярные афферентные волокна и нейроны латерального вестибулярного ядра. Их аксонов идут в спинной мозг в составе вестибулоспинального тракта. Рефлекторная дуга включает в себя небольшое число последовательно включенных нервных элементов, что обеспечивает коррекцию позы при вестибулярных раздражениях благодаря моноинаптичеким связям вестибулоспинальных волокон с мотонейронами мышц-разгибателей и торможению мышц-сгибателей. 2) Статокинетические направлены на сохранение позы и ориентацию в пространстве при изменении скорости движения. Активация вестибулярного аппарата возбуждает и вегетативные центры, в том.ч ядра блуждающего нерва. ХЗ вообще что это такое: Продолговатый мозг организует и реализует ряд защитных рефлексов: рвоты, чиханья, кашля, слезоотделения, смыкания век. Эти рефлексы реализуются благодаря тому, что информация о раздражении рецепторов слизистой оболочки глаза, полости рта, гортани, носоглотки через чувствительные ветви тройничного и языкоглоточного нервов попадает в ядра продолговатого мозга. Из него идут команды к двигательным ядрам тройничного, блуждающего, лицевого, языкоглоточного, добавочного или подъязычного нервов, в результате чего реализуется тот или иной защитный рефлекс. Точно так же за счет последовательного включения мышечных групп головы, шеи, грудной клетки и диафрагмы организуются рефлексы пищевого поведения: сосания, жевания, глотания. Продолговатый мозг также организует рефлексы поддержания позы: сигналы от рецепторов преддверия улитки и полукружных каналов поступают в верхнее вестибулярное ядро (ядро Бехтерева) переработанная здесь информация оценки необходимости изменения позы посылается к латеральному (ядру Дейтерса) и медиальному (ядро Швальбе) вестибулярным ядрам, которые участвуют в определении того, какие мышечные системы, сегменты спинного мозга должны принять участие в изменении позы от данных ядер информация направляется по вестибулоспинальному пути к передним рогам соответствующих сегментов спинного мозга, иннервирующих те мышцы, участие которых в изменении позы необходимо в данный момент. Изменение позы осуществляется за счёт: 1) статические рефлексы - регулируют тонус скелетных мышц с целью удержание определенного положения тела 2) статокинетические рефлексы- нормализуют положение тела и его позу при движении и ускорении Для реализации этих рефлексов необходима рефлексия положения и выпрямления. Рефлексия положения - обеспечивает изменение тонуса мышц при изменении положения головы относительно тела Рефлексия выпрямления - обеспечивает перераспределение тонуса мышц восстанавливающих естественное положение для данного вида позы. При ускорении направленном вверх, увеличивается тонус мышц разгибателей, если вниз, то сгибателей. Если ускорение направленно вперед, то увеличивается тонус мышц разгибателей, если назад, то сгибателей. Вегетативные рефлексы продолговатого мозга реализуются через расположенные в нём ядра блуждающих нервов, которые получают информацию о состоянии деятельности сердца, сосудов, пищеварительного тракта, лёгких, пищеварительных желез и др. В ответ на эту информацию ядра организуют двигательную и секреторную реакции данных органов. Возбуждение ядер блуждающего нерва вызывает усиление сокращения ГМК мышц желудка, кишечника, желчного пузыря и одновременно расслабление сфинктеров этих органов. При этом замедляется и ослабляется работа сердца, уменьшается просвет бронхов. Деятельность ядер блуждающих нервов проявляется также в усилении секреции бронхиальных, желудочных, кишечных желёз, в возбуждении поджелудочной железы, секреторных клеток печени. В средних отделах ретикулярной формации находятся нейроны, образующие ретикулоспинальный путь, оказывающий тормозное влияние на мотонейроны спинного, мозга. На дне 1V желудочка расположены нейроны «голубого-пятна». Их медиатором является норадреналин. Эти нейроны вызывают активацию ретикулоспинального пути в фазу «быстрого» сна, что приводит к торможению спинальных рефлексов и снижению мышечного тонуса. Мост располагается выше продолговатого мозга и выполняет сенсорные, проводниковые, двигательные, интегративные рефлекторные функции. Ретикулярная формация моста является продолжением ретикулярной формации продолговатого мозга и началом аналогичной системы среднего мозга. Аксоны нейронов ретикулярной формации моста идут в мозжечок, спинной мозг (ретикулоспинальный путь). Последние активируют нейроны спинного мозга. Ретикулярная формация моста влияет на кору большого мозга, вызывая пробуждение или сонное состояние. В ретикулярной формации моста находятся две группы ядер, которые относятся к общему дыхательному центру. Один центр активирует центр вдоха продолговатого мозга, другой — центр выдоха. |