|
|
Электронный атлас. Тема 03 Цитоплазма. Тема Цитоплазма Общий обзор
В данном процессе можно различить три основные стадии:
синтез, видимо, как мембраносвязанными, так и свободными рибосомами новых пероксисомных ферментов,
проникновение этих макромолекул (благодаря наличию у них определённой трипептидной "метки") в уже существующие пероксисомы, что приводит к увеличению объёма последних;
отпочковывание от увеличенных пероксисом новых органелл.
|

3.3. Рибосомы и митохондрии
3.1. Рибосомы
3.3.1.1. Классификация рибосом
|
Типы рибосом
|
В клетках животных различают два типа рибосом:
цитоплазматические рибосомы, которые содержатся в цитозоле и значительно преобладают по числу,
и митохондриальные рибосомы – содержатся в митохондриях, несколько отличаясь своим составом и размером (более мелким).
Далее в этом разделе речь будет идти, в основном, о цитоплазматических рибосомах.
|
Подтипы
цитоплаз-
матичес-
ких рибосом
|
а) Как мы уже знаем, рибосомы цитоплазмы подразделяются на два подтипа:
свободныерибосомы – находятся в гиалоплазме, не будучи связанными с мембранами ЭПС;
|
мембрано-
связанные рибосомы – фиксированы на наружной (обращённой к гиалоплазме) поверхности ЭПС.
|
|
б) В обоих случаях это непременно функционирующие рибосомы, причём находящиеся, как правило, в составе полисом.
|
3.3.1.2. Строение рибосомы
|
Компо-
ненты
|
а) В собранном виде цитоплазматическая рибосома состоит из двух субъединиц следующего состава:
-малая субъединица –
одна длинная цепь рРНК (около 2000 нуклеотидов, константа седиментации – 18S),
с которой связано примерно 30 молекул рибосомальных белков;
- большая субъединица –
ещё более длинная цепь рРНК ( 4000 нукл., 28S),
с которой связано 2 короткие цепи РНК (5,8S и 5S)
и около 45 молекул белков.
б) В итоге каждая субъединица представляет собой свёрнутый рибонуклеопротеидный тяж, имеющий несколько функциональных центров.
|
Место образо-
вания рРНК и белков
|
а) Рибосомные РНК (рРНК) образуются в ядрышках(о чём подробнее будет сказано в следующей теме).
б) Там же, в ядрышках, формируются и сами субъединицы рибосомы, которые затем перемещаются из ядра в цитоплазму.
в) И в неработающем состоянии субъединицы рибосом так и остаются диссоциированными друг от друга.
|
3.3.1.3. Функционирование рибосом
I. Участие рибосом в трансляции
|
Суть трансля-
ции
|
а) Основная функция рибосом состоит в том, что с их помощью осуществляется трансляция,т.е.
поочерёдное включение аминокислот в строящуюся пептидную цепь в соответствии с последовательностью кодонов в матричной РНК (мРНК).
б) В трансляции различают 3 этапа:
инициацию, элонгацию и терминацию.
|
1 Инициа-
ция
|
Инициация трансляции - это сборка функционально активного комплекса:
с определёнными центрами малой субъединицы последовательно связываются:
начальный участок мРНК,
инициирующая транспортная РНК (тРНК), нагруженная первой аминокислотой будущей пептидной цепи,
и, наконец, большая субъединица рибосомы.
|
2. Элонгация и образова-
ние полисом
|
1. а) Собранная рибосома начинает постепенно перемещаться вдоль мРНК, осуществляя процесс трансляции, т.е. удлиняя строящуюся пептидную цепь.
б) Данная стадия называется элонгацией (от elongation – удлинение).
2. а) Когда рибосома удалится на достаточное расстояние от начального участка мРНК,
с этим участком связываются субъединицы другой рибосомы – так что одну цепь мРНК начинают транслировать сразу две рибосомы.
б) И так далее: с цепью мРНК постепенно связывается несколько рибосом, что приводит к образованию полисомы.
|
Термина-
ция
|
а) Когда рибосома заканчивает трансляцию мРНК (т.е. доходит до конца кодирующей части мРНК), образуемый с её участием комплекс разрушается:
рибосома диссоциирует от мРНК, вновь распадаясь на отдельные субъединицы;
одновременно высвобождается и готовая пептидная цепь.
б) Этот процесс обозначается как терминация трансляции.
|
II. Полисомы
Задержим своё внимание на полисомах.
|
Общая характе-
ристика
|
В полисоме рибосомы
находятся на примерно равном расстоянии друг от друга,
движутся по мРНК в одном направлении,
и каждая синтезирует “свою” пептидную цепь.
|
Смена состава рибосом
|
Состав рибосом в полисоме постепенно сменяется:
рибосомы, заканчивающие трансляцию мРНК, покидают полисому,
а с начальным участком мРНК связываются субъединицы иных рибосом (до тех пор, пока мРНК не будет разрушена специальными РНКазами).
|
О понятии "свобод-
ные рибосомы"
|
а) Таким образом, термины “свободные” и “мембраносвязанные” следовало бы применять не к отдельным рибосомам, а к полисомам.
б) Действительно, как уже отмечалось, рибосомы в свободном состоянии практически не бывают:
когда они не транслируют какую-либо мРНК, они диссоциированы на отдельные субъединицы.
|
Мембрано-
связанные полисомы
|
Что же касается полисом, то, согласно п. 3.2.1,
мембраносвязанными они становятся при образовании экспортных, мембранных, лизосомных и ряда пероксисомных белков,
причём с мембраной связывается каждая рибосома полисомы – как только в синтезируемой ею пептидной цепи образуется сигнальная последовательность (СП).
|
Свободные полисомы
|
а) Свободные же полисомы синтезируют белки, которые
либо остаются в гиалоплазме,
либо переходят в состав тех или иных клеточных структур (ядрa, митохондрий, цитоскелета, пероксисом).
б) Содержание свободных полисом (а в их составе – и рибосом) особенно велико в быстро растущих клетках.
|
Общее представ-
ление
|
а) После образования на рибосомах пептидной цепи должно произойти сворачивание белка в соответствующую трёхмерную структуру.
Данный процесс обозначается как фолдинг.
б) Конкретный вид трёхмерной структуры белка полностью определяется его первичной структурой (т.е. последовательностью аминокислот).
в) Но, видимо, во многих случаях достижение белком правильной трёхмерной структуры значительно ускоряют специальные белки:
традиционные ферменты и
т.н. молекулярные шапероны.
|
Ферменты фолдинга
|
а) Один из ферментов фолдинга катализирует разрыв и образование в белке дисульфидных связей.
б) Это ускоряет разрыв "неправильных" и замыкание "правильных" дисульфидных связей.
|
Шапероны
|
а) Что касается шаперонов, то они, видимо, связываются с пептидной цепью белка ещё на рибосоме - до завершения синтеза всей цепи.
б) Тем самым они препятствуют "неправильному" сворачиванию уже образованного фрагмента цепи.
в) В ряде случаев связь с шаперонами сохраняется некоторое время и по окончании синтеза белка на рибосоме.
Например, в таком виде траспортируются белки митохондрий от цитоплазматических рибосом в сами митохондрии.
г) После диссоциации шаперонов белок получает возможность быстро принять правильную трёхмерную структуру.
|
Тепловой стресс
|
а) При тепловом стрессе белки теряют свою нативную (правильную) конфигурацию.
б) В этом случае усиливается синтез шаперонов (которые ещё называются "белками теплового шока").
в) Последние
способствуют полному разворачиванию повреждённых белков и
затем диссоциируют.
г) После этого белок вновь может вернуться к нативной конфигурации.
|
3.3.1.5. Цитохимическое обнаружение рибосом по РНК
|
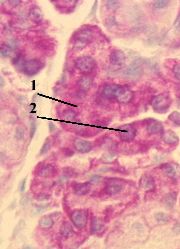
5. Препарат - РНК в цитоплазме и ядрышке клеток (поджелудочная железа). Окраска по Браше (метиловым зелёным - пиронином).
1. Применённый метод окраски (по Браше) выявляет РНК, которая окрашивается в малиновый цвет.
2. На препарате РНК обнаруживается в цитоплазме (1) и ядрышках (2) клеток.
3. а) Основная часть этой РНК и там, и там представлена рибосомными РНК.
б) Доля матричных и транспортных РНК в общем пуле клеточной РНК сравнительно невелика.
|
Полный размер
|
3.3.2. Митохондрии
3.3.2.1. Строение
I. Общие сведения
|
Две мембраны
|
а) Митохондрии относятся к мембранным структурам цитоплазмы.
б) Однако, как уже отмечалось, они имеют не одну, а две мембраны.
|
Наружная мембрана
|
Наружная мембрана (1)
содержит широкие гидрофильные каналы и потому
хорошо проницаема для многих веществ (кроме высокомолекулярных).
|
Внутрен-
няя мембрана
|
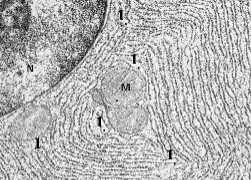
а) Внутренняя мембрана (2) образует многочисленные впячивания (кристы) (3) в матрикс (4) митохондрии.
б) На кристах имеются грибовидные выросты – оксисомы, или F1-частицы.
в)В кристы и оксисомы встроены ферменты дыхательной цепи и синтеза АТФ.
|
Электронная
микрофотография –
митохондрия.

Полный размер
|
Матрикс
|
Внутреннее пространство митохондрий (между кристами) заполнено матриксом, состав которого станет ясен из последующего изложения.
|
II. Вариабельность структуры митохондрий
|
1. а) Форма митохондрий варьирует от почти сферической до очень вытянутой.
б) В некоторых клетках митохондрии имеют ещё более сложную форму: например, образуют разветвления.
2. Различаются также количество и форма крист: это демонстрируется следующими тремя примерами.
|
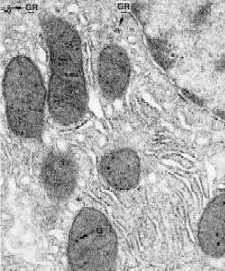
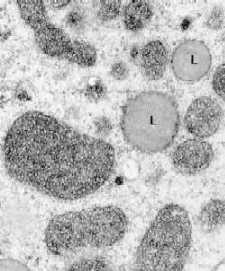
Электронные микрофотографии – митохондрии в разных тканях.
|
а) Мышечное волокно
|
б) Клетка печени
|
в) Клетка коры надпочечников
|

Полный размер
|
Полный размер
|
Полный размер
|
В мышечных волокнах, где потребности в энергии особенно велики, в митохондриях
крист очень много
и они имеют пластинчатую (ламинарную) структуру.
|
В клетках печени количество крист в митохондриях
значительно меньше.
|
Наконец, в клетках коры надпочечников кристы
имеют тубулярную структуру и
выглядят на срезе как мелкие везикулы.
|
3.3.2.2. Система автономного синтеза белков в митохондриях
|
Компо-
ненты системы
|
Митохондрии отличаются от прочих органелл ещё двумя особенностями: они содержат в своём матриксе
а) собственную ДНК (мтДНК) –
от 1 до 50 небольших одинаковых циклическихмолекул, включающих по 37 генов,
б) и собственные рибосомы, которые
по размеру несколько меньше цитоплазматических рибосом и
на электронных микрофотографиях выглядят как мелкие гранулы
|
Роль системы
|
а) Данная система автономного синтеза белков обеспечивает образование
всего лишь 5 % митохондриальных белков.
б) Остальные белки митохондрий
кодируются молекулами ядерной ДНК
и синтезируются цитоплазматическими рибосомами.
|
Проис-
хождение
мито-
хондрий
|
а) Структура мтДНК и рибосом сближает митохондрии с бактериями: у последних тоже –
циклическая ДНК и
относительно небольшие рибосомы.
б) Поэтому возможно, что в эволюции митохондрии появились как
результат симбиоза древних бактерий с эукариотическими клетками.
|
Мутации мтДНК
|
а) МтДНК находится в среде, где очень активно идут окислительные процессы, побочным продуктом которых являются свободные радикалы.
б) К тому же эта ДНК не связана с многочисленными белками.
в) Поэтому
частота мутаций мтДНК в 10 раз выше, чем ядерной ДНК.
|
Наследо-
вание мтДНК
|
а) Но имеют значение мутации только материнской мтДНК(в женских половых клетках), поскольку
лишь эта мтДНК передаётся потомству.
б)Митохондрии же сперматозоида в яйцеклетку не попадают.
|
Реплика-
ция мтДНК
|
Наконец, отметим, что репликация (удвоение) мтДНК происходит в любой фазе клеточного цикла (исключая само митотическое деление).
|
3.3.2.3. Функции митохондрий
| |
|
|
 Скачать 1.22 Mb.
Скачать 1.22 Mb.