Тема 3.
Цитоплазма

3.1. Общий обзор
3.1.1. Состав цитоплазмы
В цитоплазме клеток обычно содержатся следующие компоненты:
гиалоплазма (другое название – цитозоль),
органеллы и
включения.
3.1.1.1. Гиалоплазма
Опреде-
ление
|
Гиалоплазма - это матрикс цитоплазмы, в котором находятся органеллы и включения.
|
Состав
|
Гиалоплазма представляет собой водный раствор
неорганических ионов,
органических метаболитов,
биополимеров (белков, полисахаридов, транспортных РНК и т.д.).
|
Структу-
рирован-
ность
|
а) Некоторые макромолекулы могут объединяться (путём самосборки) в те или иные комплексы.
б) Таким образом, и гиалоплазма, видимо, в определённой степени структурирована.
|
Консис-
тенция
|
По консистенции она приближается к гелю.
|
3.1.1.2. Органеллы
Опреде-
ление
|
Органеллы - это такие морфологически различимые структуры цитоплазмы, которые
обязательно должны присутствовать в данной клетке, выполняя в ней определённые функции.
|
Класси-
фикация по распростра-
нённости
|
а) По своей распространённости органеллы делятся на две группы:
органеллы общего значения – содержатся практически во всех клетках,
и органеллы специального значения – имеются только в клетках (или надклеточных структурах) какого-то определённого вида, обеспечивая выполнение их специфических функций.
б) Примеры органелл второй группы –
миофибриллы в мышечных волокнах и клетках,
а также реснички в покровных клетках дыхательных путей.
|
Класси-
фикация по строению
|
По строению органеллы тоже подразделяются на два типа:
мембранные органеллы – отграничены собственной мембраной от окружающей гиалоплазмы, т.е. являются замкнутыми компартментами (отсеками);
и немембранные органеллы – структуры, не окружённые мембраной.
|
3.1.1.3. Включения
Опреде-
ление
|
Включения – это необязательные компоненты цитоплазмы данной клетки (тоже различимые морфологически):
они возникают и исчезают в зависимости от состояния клетки или организма в целом.
|
Виды включений
|
Включения можно классифицировать различными способами:
а) по функциональной роли –
резервные (в т.ч. трофические),
транспортные (в т.ч. секреторные),
светозащитные,
балластные и т.д.;
б) по физическим свойствам –
пигментные и
непигментные;
в) по происхождению в клетке –
экзогенные и
эндогенные.
|
Резервные (и трофи-
ческие) включения
|
1. К резервным включениям относятся, в частности,
гранулы гемосидерина в клетках печени (содержат запасы железа в комплексе с белком ферритином),
гранулы гликогена вновь в клетках печени, а также в мышечных волокнах,
капельки нейтрального жира в жировых и некоторых других клетках.
2. а) Поскольку гликоген и жиры являются питательными веществами, их включения в клетках обозначаются как трофические.
б) Но, по существу, это такое же резервирование необходимых веществ, что и отложение железа в составе гемосидерина.
|
Включения транс-
портного типа
|
Включения транспортного типа образуются в результате мультимолекулярного переноса веществ в клетку или из клетки, это
продукты эндоцитоза - пиноцитозные пузырьки и фагосомы,
продукты экзоцитоза - секреторные и экскреторные мембранные пузырьки.
|
Свето-
защитные включения
|
Светозащитные включения – это гранулы меланина, присутствующие в определённых клетках. Данные включения являются
пигментными, т.е. поглощают свет, и на этом свойстве основана их светозащитная функция.
|
Другие пигмент-
ные включения
|
1. В клетках могут присутствовать и другие пигментные включения – с совершенно иной функцией. Примеры:
уже упоминавшиеся гранулы гемосидерина (функция – резервирование железа),
балластный пигмент липофусцин (см. п. 3.2.4) и т.д.
2. Как пигментное включение иногда рассматривают и гемоглобин в эритроцитах (хотя его нельзя считать необязательным компонентом этих клеток).
3. Таким образом, способность включения поглощать свет далеко не всегда определяет функцию этого включения.
|
3.1.2. Демонстрация включений
3.1.2.1. Включения гликогена
1,а-б. Препарат - включения гликогена в клетках печени. Окраска по Бесту.
|
а) (Малое увеличение)
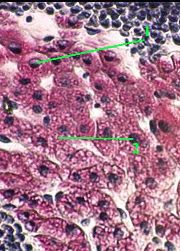
Полный размер
|
б) (Большое увеличение)
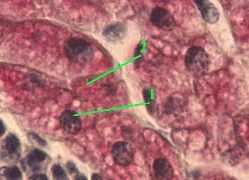
Полный размер
|
1. На снимках видны клетки печени (гепатоциты) с ядром (1) фиолетового цвета.
2. В цитоплазме - многочисленные глыбки гликогена (2), окрашенные в ярко-красный цвет.
|
3.1.2.2. Жировые включения
2. Препарат - жировые включения в клетках печени аксолотля. Фиксация осмиевой кислотой, окраска кармином.
|
1. а) При фиксации осмиевой кислотой жировые капли, будучи осмиофильными, поглощают фиксатор.
б) Поэтому при последующей окраске кармином
прочие структуры приобретают красноватый оттенок,
а жировые капли, содержащие соединения осмия, сохраняют чёрный цвет.
|
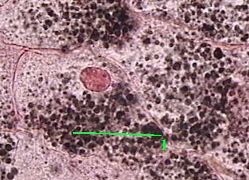
Полный размер
|
2. В соответствии с этим, в цитоплазме печеночных клеток мы видим чёрные жировые включения (1) разного размера.
|
3.1.3. Классификация органелл цитоплазмы
Далее речь будет идти только об органеллах. Вот их краткий перечень.
3.1.3.1. Мембранные органеллы
I. Вакуолярная система
За исключением митохондрий, все мембранные органеллы цитоплазмы объединяются в т.н. вакуолярную систему. В последнюю входят следующие органеллы.
1. Эндоплазма-
тическая сеть (ЭПС)
|
а) Другое название - эндоплазматический ретикулум.
б) Это совокупность плоских мембранных мешков (цистерн), вакуолей и трубочек.
|
2. Комплекс (аппарат) Гольджи
|
- несколько скоплений плоских мембранных цистерн, от которых отшнуровываются мелкие пузырьки.
|
3. Лизосомы
|
- мембранные пузырьки, содержащие ферменты гидролиза биополимеров (протеазы, нуклеазы, гликозидазы, липазы и т.д.).
|
4. Перокси-
сомы
|
- мембранные пузырьки, содержащие
ферменты, которые нейтрализуют пероксид водорода, а также
оксидазы - ферменты окисления субстратов непосредственно кислородом.
|
II. Митохондрии
Это органеллы, отграниченные (как и ядро) двумя мембранами, из которых
внутренняя образует многочисленные впячивания (кристы) внутрь митохондрии.
|
3.1.3.2. Немембранные органеллы
Данные органеллы тоже можно подразделить на две неравные группы: глобулярные и фибриллярные.
I. Глобулярные органеллы
Сюда относятся только рибосомы - многочисленные небольшие частицы, состоящие из
двух субъединиц рибонуклеопротеидной природы.
|
II. Фибриллярные органеллы
Сюда входят:
сократительные структуры,
элементы цитоскелета и
производные элементов цитоскелета.
А.Сократительные структуры – это, как уже упоминалось,
миофибриллы и миофиламенты.
|
Б. Элементы цитоскелета.
|
1. Микро-
филаменты
(МФ)
|
– нити (d = 5 7 нм) из белка актина, пронизывающие гиалоплазму в тангенциальном направлении (вдоль длинной оси клетки).
|
2. Микро-
трубочки
(МТ)
|
– полые трубки (d = 24 нм) из белка тубулина, имеющие, в основном, радиальную ориентацию в клетке.
|
3. Промежу-
точные
филаменты
|
– нити промежуточного диаметра (d = 10 нм),
причём, в клетках разных тканей они образованы разными белками.
|
В. Производные элементов цитоскелета
|
1,а. Каркас микро-
ворсинок
|
Каркас микроворсинок образован параллельными пучками микрофиламентов (см. п. 2.4.1)
|
2,а. Центриоли
|
а) Центриоль представляет собой полый цилиндр, образованный микротрубочками.
б) В клетке обычно содержится пара центриолей (диплосома), причём она
входит в состав клеточного центра– органоида общего значения.
|
2,б. Аксонема
|
Аксонема ("осевая нить")
тоже (как центриоли) состоит из микротрубочек и
служит цитоскелетом ресничек и жгутиков.
|
3.1.4. Схема строения клетки
1. Многие из вышеперечисленных структур показаны на данной схеме.
а) Компоненты вакуолярной системы цитоплазмы
эндоплазматическая сеть (1),
комплекс Гольджи (2),
лизосомы (3).
б) Другие компоненты цитоплазмы:
митохондрии (4),
рибосомы (5), центриоль (6).
в) Ядро (7) и в нём -
ядерная оболочка (8) и ядрышко (9).
|
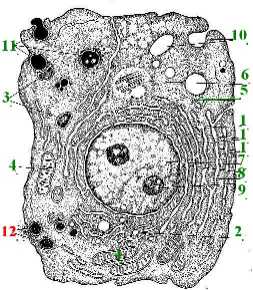
Полный размер
|
2. Кроме того, мы здесь видим структуры, участвующие в экзо- и эндоцитозе, -
пиноцитозные пузырьки (10),
фагосомные вакуоли (11),
секреторные вакуоли (12).
|
Теперь рассмотрим перечисленные в таблице структуры подробнее.
3.2. Вакуолярная система цитоплазмы
Эндоплазматическая сеть (ЭПС) делится на два типа -
гранулярную и
агранулярную (или гладкую).
3.2.1. Гранулярная ЭПС
3.2.1.1. Компоненты
Общий вид
|
Как видно на микрофотографии, ЭПС (1), действительно, представляет собой совокупность плоских мешков (цистерн), вакуолей и трубочек, заполняющих значительную часть цитоплазмы.
|
Электронная
микрофотография -
гранулярная
эндоплазмати-
ческая сеть.
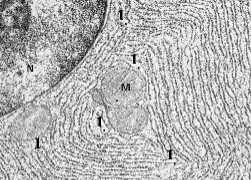
Полный размер
|
Единство ЭПС
|
а) На самом деле практически все эти структуры (за исключением отдельных пузырьков) – части единого непрерывного компартмента (отсека), отграниченного мембраной от гиалоплазмы.
|
б) И отдельными (не сообщающимися друг с другом) они кажутся только на срезе.
|
Ключе-
вая особен-
ность
|
а) Особенность гранулярной ЭПС состоит в том, что со стороны гиалоплазмы мембранная сеть покрыта мелкими гранулами - рибосомами.
б) В связи с этим, иногда используют другой термин - шероховатый ретикулум.
|
3.2.1.2. Функции гранулярной ЭПС
Спектр синтези-
руемых белков
|
На рибосомах гранулярной ЭПС происходит синтез белков, причём таких, которые затем
либо выводятся из клетки (экспортные белки),
либо входят в состав определённых мембранных структур (собственно мембран, лизосом, пероксисом).
|
Меха-
низм связыва-
ния рибосом с ЭПС
|
а) Пептидные цепи указанных белков (точнее, их предшественников) начинаются с т.н. сигнальной последовательности (СП).
б) Эта СП, как только образуется в ходе трансляции,
связывается с мембраной ЭПС (в месте нахождения специальных докинг-белков) и
проникает своим начальным концом в полость ЭПС, формируя трансмембранный канал.
в) Все остальные части синтезируемой пептидной цепи, следуя за СП по каналу, тоже оказываются в полости ЭПС.
|
Оконча-
ние
синтеза пептид-
ной цепи
|
а) По окончании синтеза пептидной цепи она теряет связь с образовавшей её рибосомой, благодаря чему
рибосома перестаёт быть мембраносвязанной,
а трансмембранный канал исчезает.
б) Сам же белок (пептидная цепь) остаётся по другую сторону мембраны ЭПС – в просвете ЭПС.
в) От белка отщепляется СП.
|
Фолдинг белка
|
Здесь же (в просвете ЭПС) происходит фолдинг белка –
сворачивание его пептидной цепи в правильную трёхмерную структуру.
(Подробней о фолдинге будет сказано в п. 3.3.1.2.)
|
Начало модифи-
кации белка
|
а) В просвете ЭПС также начинается модификация белка.
Чаще всего это проявляется в том, что к пептидной цепи белка (будущего гликопротеина)
присоединяются небольшие разветвлённые олигосахаридные цепочки.
б) Эти цепочки образуются в гиалоплазме и переносятся в просвет ЭПС с помощью специального вещества – долихолфосфата.
|
Перечень функций
|
В итоге, можно перечислить следующие функции гранулярной ЭПС:
синтез на рибосомах пептидных цепей экспортных, мембранных, лизосомных и отчасти пероксисомных белков,
фолдинг белков,
изоляция этих белков от гиалоплазмы внутри мембранных полостей и концентрирование их здесь,
начальная химическая модификация этих белков, а также
их транспорт (внутри ЭПС и с помощью отдельных пузырьков).
|
Вывод
|
а) Таким образом, наличие в клетке хорошо развитой гранулярной ЭПС свидетельствует о высокой интенсивности белкового синтеза - особенно в отношении секреторных (экспортных) белков.
б) В частности, это имеет место
в клетках, синтезирующих гормоны белковой природы.
|
3.2.2. Комплекс Гольджи
3.2.2.1. Состав комплекса
Схема - строение и функциони-
рование комплекса Гольджи.
|
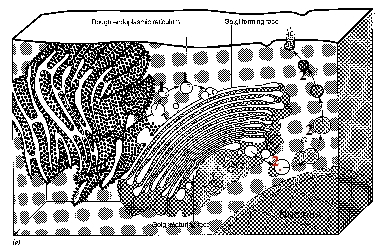
Полный размер
|
Связь с ЭПС
|
Белки, синтезированные на гранулярной ЭПС, перемещаются
по внутреннему её пространству или
в составе транспортных пузырьков (покрытых клатрином)
к одной из диктиосом (1) комплекса Гольджи.
|
Состав комп-
лекса
|
а) Диктиосома – это скопление 5-10 плоских мембранных цистерн, лежащих параллельно друг другу. Иногда в понятие диктиосомы включают также близлежащие пузырьки и вакуоли.
б) В клетке может быть много диктиосом, соединённых с ЭПС и друг с другом цистернами и трубочками.
Вся совокупность диктиосом и составляет комплекс Гольджи.
|
Поляр-
ность диктио-
сомы
|
а) В каждой диктиосоме различают две части:
проксимальную (cis-), обращённую к ЭПС,
и дистальную (trans-), расположенную с противоположной стороны.
б) Таким образом, белки, транспортированные из ЭПС, оказываются вначале в проксимальной части той или иной диктиосомы.
|
3.2.2.2. Функционирование комплекса Гольджи
|
 Скачать 1.22 Mb.
Скачать 1.22 Mb.