Тема роль ствола мозга, базальных ганглиев и коры больших полушарий в регуляциидвигательныхфункцийорганизма
 Скачать 168.97 Kb. Скачать 168.97 Kb.
|

1 2 Тема 4. РОЛЬ СТВОЛА МОЗГА, БАЗАЛЬНЫХ ГАНГЛИЕВ И КОРЫ БОЛЬШИХ ПОЛУШАРИЙ В РЕГУЛЯЦИИДВИГАТЕЛЬНЫХФУНКЦИЙОРГАНИЗМА 1. Функции продолговатого мозга. Роль заднего мозга (вестибулярных ядер и ретикулярной формации) в обеспечении антигравитационной позы. Восходящие и нисходящие пути. 2. Функции двигательных ядер среднего мозга. Децеребрационная ригидность. Ориентировочный рефлекс. 3. Двигательные рефлексы ствола мозга: статические и стато-кинетические. 4. Функции ретикулярной формации ствола мозга. 5. Морфо-функциональная характеристика мозжечка. Мозжечковые пробы. Симптомы поражения мозжечка. 6. Функции базальных ядер, последствия нарушения их функционирования у человека (гипер- и гипокинезы, гипер- и гипотонус). 7. Структурно-функциональная организация коры больших полушарий (слои, колонки). Локализация функций в коре больших полушарий. Роль коры головного мозга в регуляции двигательной активности организма. Продолговатый мозг – является непосредственным продолжением спинного мозга. Он сочетает в себе черты строения спинного мозга и начального отдела головного мозга. На его передней поверхности по средней линии проходит передняя срединная щель, являющаяся продолжением одноименной борозды спинного мозга. По бокам от щели находятся пирамиды, которые продолжаются в передние канатики спинного мозга. Пирамиды состоят из пучков нервных волокон, которые в борозде перекрещиваются с такими же волокнами противоположной стороны. Латеральнее пирамид с двух сторон находятся возвышения – оливы. На задней поверхности продолговатого мозга проходит задняя (дорсальная) срединная борозда, являющаяся продолжением одноименной борозды спинного мозга. По бокам от борозды лежат задние канатики. В них проходят восходящие пути спинного мозга. По направлению кверху задние канатики расходятся в стороны и идут к мозжечку. Функции продолговатого мозга. 1. Рефлекторная функция связана с центрами находящимися в продолговатом мозге. В продолговатом мозге расположены следующие центры: 1) Дыхательный центр, обеспечивающий вентиляцию легких; 2) Пищевой центр, регулирующий сосание, глотание, отделение пищеварительного сока (слюноотделения, желудочного и поджелудочного соков); 3) Сердечно-сосудистый центр – регулирующий деятельность сердца и кровеносных сосудов. 4) Центр защитных рефлексов – это мигание, слюноотделение, чихание, кашель, рвота. 5) Центр лабиринтных рефлексов, осуществляющий распределение мышечного тонуса между отдельными группами мышц и установочные рефлексы позы. Задний мозг включает в себя продолговатый мозг и варолиев мост. Основными структурно-функциональными образованиями заднего мозга являются: 1) ядра Голля и Бурдаха, 2) ядра 5-12 пар черепно-мозговых нервов, 3) вестибулярные ядра Дейтерса, Бехтерева, Швальбе, 4) ретикулярная формация, 5) жизненно важные центры, 6) центр пищеварения, 7) центры защитных рефлексов. Ядра Голля и Бурдаха обеспечивают проведение кожно-механической чувствительности. Ядра 5-12 пар черепно-мозговых нервов непосредственно регулируют деятельность иннервируемых органов и тканей. Вестибулярные ядра и ретикулярная формация заднего мозга обеспечивают регуляцию мышечного тонуса. Жизненно важные центры заднего мозга участвуют в регуляции основных необходимых для поддержания жизнедеятельности процессов – дыхания и кровообращения. К ним относят: 1) дыхательный, 2) сосудодвигательный, 3) сердечный. Повреждение этих центров не совместимо с жизнью. Пищеварительный центр заднего мозга регулирует моторную, секреторную и всасывательную функции пищеварительного тракта. Центры защитных рефлексов - кашля, чихания, мигания и рвоты, обеспечивают защиту организма от повреждающих факторов. Выделяют три основных общих функции заднего мозга: 1) рефлекторная, 2) проводниковая, 3) трофическая. Проводниковая функция заднего мозга заключается в обеспечении двусторонних связей между ниже и вышерасположенными отделами ЦНС. Трофическая функция заднего мозга заключается в питании и регуляции метаболизма органов и тканей, иннервируемых нейронами заднего мозга. Рефлекторная функция заднего мозга может быть сегментарной и надсегментарной. Рефлекторная сегментарная функция заднего мозга заключается в непосредственном влиянии эфферентных нейронов заднего мозга на эффекторы при раздражении рецепторов рефлексогенной зоны. Рефлекторная надсегментарная функция заднего мозга заключается в регуляции сложнокоординированных рефлекторных актов через нижерасположенные отделы ЦНС. Она связана с деятельностью вестибулярных ядер, ретикулярной формации и центров регуляции жизненно важных функций - дыхательного и сосудодвигательного. 2.3.3. Функции ретикулярной формации Ретикулярная формация (РФ) - это совокупность нейронов, диффузно расположенных в центральной части заднего, среднего и промежуточного мозга. Ретикулярная формация оказывает как восходящее, так и нисходящее влияние на пограничные отделы ЦНС. Восходящее влияние РФ заключается в неспецифическом активирующем воздействии на всю кору больших полушарий, что обеспечивает поддержание бодрствующего состояния головного мозга. Поэтому наиболее ярким проявлением при блокаде ретикулярной формации будет беспробудный сон или коматозное состояние. Нисходящее влияние РФ проявляется как в активирующем, так и в тормозящем ее действии на спинальные нервные центры. Нейроны ретикулярной формации характеризуются: 1) высокой возбудимостью, 2) постоянной тонической активностью, 3) высокой чувствительностью к гуморальным раздражителям. Восходящие и нисходящие пути спинного мозга Проводниковая функция спинного мозга заключается в том, что через него проходят восходящие и нисходящие пути. К восходящим путям относятся: система задних канатиков (нежный и клиновидный пучки), являющихся проводниками кожно-механической чувствительности в продолговатый мозг; спиноталамические пути, по которым импульсы от рецепторов поступают к таламусу; спиномозжечковые пути (дорсальный и вентральный) участвуют в проведении импульсации, поступающей от кожных рецепторов и проприорецепторов в мозжечок. К нисходящим путям относятся: пирамидный, или кортикоспинальный, путь; экстрапирамидные пути, включающие руброспинальный, ретикулоспинальный, вестибулоспинальный тракты. Эти нисходящие пути обеспечивают влияние высших отделов центральной нервной системы на функцию скелетных мышц. Классификация восходящих путей спинного мозга 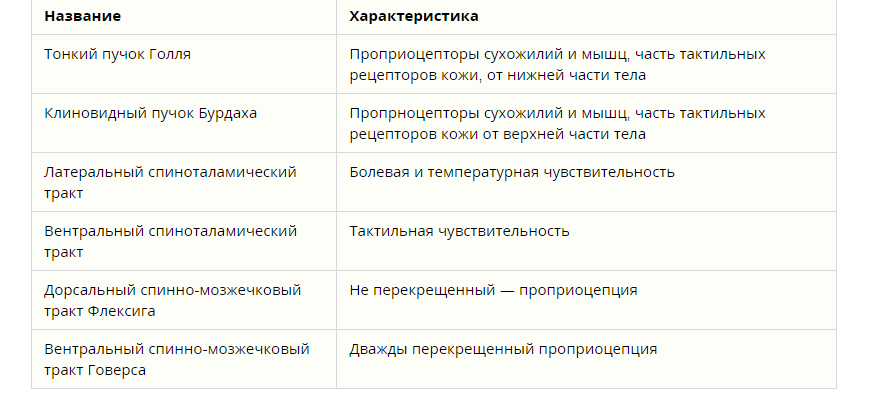 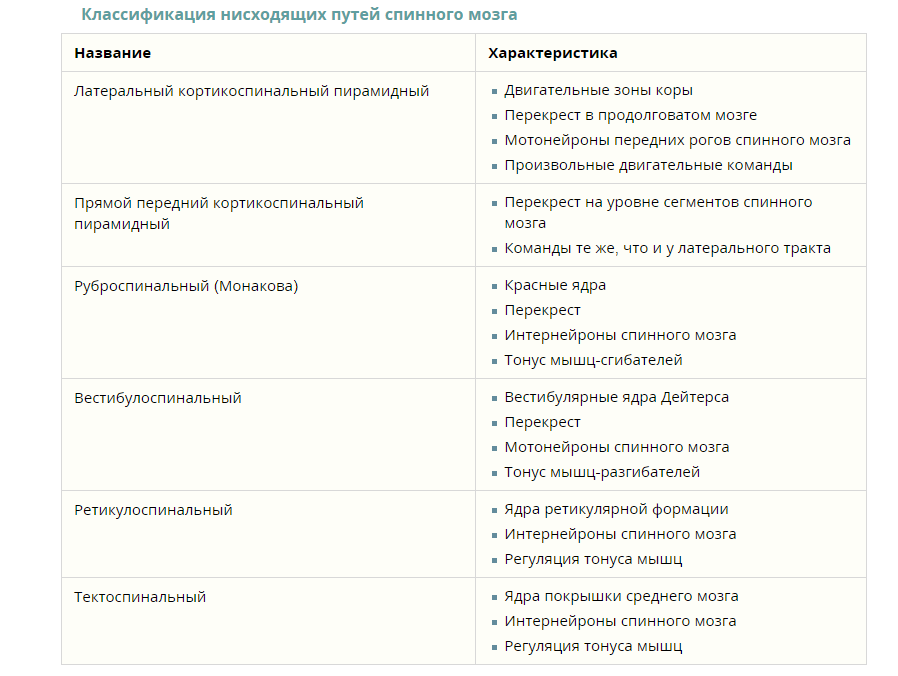 Функции продолговатого мозга. Роль заднего мозга (вестибулярных ядер и ретикулярной формации) в обеспечении антигравитационной позы. В продолговатом мозге отсутствует сегментарное строение, он имеет длину около 25 мм, серое вещество образует отдельные скопления нейронов – ядра. Продолговатый мозг выполняет собственные и проводниковые функции. Собственные функции продолговатого мозга. В продолговатом мозге расположены ядра VIII, IX, X, XI и XII пар черепно-мозговых нервов. VIII пара – преддверно-улитковый нерв (n. vestibulocochlearis). Волокна улитки, или собственно слухового нерва, являются началом слуховых путей, идущих от спирального ганглия улитки (слуховые функции). IX пара – языкоглоточный нерв (n.glossopharyngeus). Содержит двигательные волокна (иннервация мышц глотки и полости рта), отвечающие за поднимание глотки и гортани, опускание мягкого нёба и надгортанника и участвующие в рефлексе глотания, чувствительные волокна от вкусовых, тактильных, температурных, болевых, интероцептивных рецепторов задней трети языка, слизистой оболочки глотки, барабанной полости и каротидного тельца (участие в сердечно-сосудистых рефлексах) и вегетативные (парасимпатические) волокна, стимулирующие секрецию околоушной слюнной железы. Х пара – блуждающий нерв (n. vagus) – имеет три ядра. Вегетативное ядро отвечает за парасимпатическую иннервацию гортани, пищевода, сердца, желудка, тонкой кишки, пищеварительных желез. Нервные волокна чувствительного ядра образуют солитарный тракт в продолговатом мозге. Ядро получает информацию от рецепторов альвеол легких и других органов грудной и брюшной полости, участвует в качестве афферентного звена в дыхательном, глотательном и жевательном рефлексах. Двигательное ядро отвечает за сокращение мышц глотки, гортани, верхней части пищевода при глотании и дыхании, участвуя в рефлексах рвоты, чихания, кашля и в формировании голоса. ХI пара – добавочный нерв (n. accessorius) – двигательный нерв, иннервирующий грудино-ключично-сосцевидную и трапециевидную мышцы своей стороны. Сокращения этих мышц вызывают наклоны головы набок с поворотом лица в противоположную сторону, поднимание плечевого пояса и приведение лопаток к позвоночнику. При поражении нерва снижается тонус иннервируемых мышц и плечо на стороне поражения опускается, а поворот головы в сторону, противоположную поражению, затруднен. XII пара – подъязычный нерв (n. hypoglossus) – двигательный нерв, иннервирующий мышцы языка, вызывая его движения во время жевания, сосания, глотания и осуществления речи. Вегетативные функции продолговатого мозга. Продолговатый мозг содержит дыхательный центр (центр вдоха – инспираторный и центр выдоха – экспираторный). Центр вдоха расположен в основном в дорсальной части продолговатого мозга и на 95% состоит из инспираторных нейронов, аксоны которых идут в составе ретикулоспинальных путей, иннервирующих ядра диафрагмального нерва. Другая часть инспираторных нейронов, входя в состав ретикулярных вентральных ядер, контактирует с мотонейронами межреберных мышц. Нейроны центра выдоха локализуются в основном в вентральной части продолговатого мозга, включающей задние и верхние части ретикулярного гигантоклеточного ядра и ретикулярного мелкоклеточного ядра. Их аксоны иннервируют мотонейроны внутренних межреберных и брюшных мышц. В продолговатом мозге находится главный сосудодвигательный центр – регулирует тонус сосудов и уровень кровяного давления и центр сердечной деятельности – группа нейронов ядра блуждающего нерва (тормозящая) и группа нейронов, связанная со спинальными центрами (стимулирующая). Блуждающий нерв регулирует (усиливает) моторику желудочно-кишечного тракта и его секреторную деятельность. В продолговатом мозге находится центр слюноотделения, парасимпатическая часть которого обеспечивает выделение большого количества жидкой слюны, богатой неорганическими веществами, а симпатическая – небольшого количества густого белкового секрета. Продолговатый мозг отвечает за некоторые защитные рефлексы: рвоты, кашля, чихания, слезоотделения, смыкания век, а также рефлексы пищевого поведения (сосания, жевания, глотания). Продолговатый мозг реализует соматические рефлексы: рефлексы поддержания позы тела за счет статических рефлексов, регулирующих тонус мышц, удерживающих положение тела в пространстве в покое. В свою очередь статические рефлексы были разделены голландским физиологом Р. Магнусом (1924) на тонические рефлексы позы (положения тела при спокойном стоянии) и выпрямительные рефлексы. Рефлексы позы можно наблюдать при изменении положения головы по отношению к туловищу. При этом должна быть сохранена нормальная поза тела. На дне IV желудочка продолговатого мозга находятся нейроны «голубого пятна», которые выделяют медиатор норадреналин. Через ретикулоспинальный тракт эти нейроны тормозят спинальные рефлексы и снижают мышечный тонус в фазу «быстрого сна». Продолговатый мозг регулирует некоторые анализаторные функции, осуществляя первичный анализ сенсорных раздражителей (кожного, вкусового, слухового, вестибулярного). Проводниковая функция продолговатого мозга. В продолговатом мозге начинаются нисходящие пути: вестибулоспинальный, оливоспинальный, ретикулоспинальный, обеспечивающие связь между вестибулярными ядрами, оливой, ретикулярной формацией продолговатого мозга и мотонейронами спинного мозга, отвечающими за тонус и координацию мышечных реакций. Раздражение ретикулярной формации продолговатого мозга оказывает нисходящие влияния на моторную активность скелетных мышц, а также восходящие, активирующие влияния на кору больших полушарий, вызывая десинхронизацию электроэнцефалограммы. Через продолговатый мозг проходят все восходящие и нисходящие пути спинного мозга: спиноталамический, кортикоспинальный, руброспинальный. В нем заканчиваются пути из коры больших полушарий – корковоретикулярный, а также восходящие пути проприоцептивной чувствительности – тонкий пучок Голля и клиновидный пучок Бурдаха. Поэтому при повреждении одной из половин продолговатого мозга выше перекреста восходящих путей проприоцептивной чувствительности происходит нарушение чувствительности 132 и работы мышц лица и головы на стороне повреждения, в то время как на поврежденной стороне возникают нарушения кожной чувствительности и двигательные параличи туловища и конечностей. Это связано с тем, что все восходящие и нисходящие пути перекрещиваются. Ретикулярная формация Ретикулярная формация (РФ) располагается в центральной части мозгового ствола, заходя ростральным концом в таламус, а каудальным – в спинной мозг. Различные по форме и величине нейроны РФ имеют длинные дендриты и короткий аксон, хотя встречаются гигантские нейроны с длинными аксонами, образующими, например, руброспинальный и ретикулоспинальный тракты. На одной нервной клетке может заканчиваться до 40 000 синапсов, что указывает на широкие межнейрональные связи в пределах РФ. В ней был выделен целый ряд ядер и ядерных групп, отличающихся как в структурном отношении, так и выполняемыми ими функциями. Ретикулярная формация образует многочисленные как афферентные пути: спиноретикулярный, церебеллоретикулярный, корково-подкорково-ретикулярный (от коры, базальных ганглиев, гипоталамуса), от структур каждого уровня ствола мозга (от среднего мозга, варолиева моста, продолговатого мозга), так и эфферентные нисходящие: ретикулоспинальные, ретикуло-корково-подкорковые, ретикуломозжечковые, к другим структурам ствола мозга. Ретикулярная формация оказывает генерализованное, тонизирующее, активирующее влияние на передние отделы головного мозга и кору больших полушарий (восходящая активирующая система РФ), и нисходящее, контролирующее деятельность спинного мозга (нисходящая ретикулоспинальная система), которое может быть как облегчающим на многие функции организма, так и тормозным. Одним из видов тормозного влияния РФ на рефлекторную деятельность спинного мозга является «сеченовское торможение», заключающееся в угнетении спинальных рефлексов при раздражении таламической ретикулярной формации кристалликом соли Г. Мэгун показал, что локальное электрическое раздражение гигантоклеточного ядра РФ продолговатого мозга вы- 166 зывает торможение сгибательного и разгибательного рефлексов спинного мозга, а на мотонейроне возникают длительный ТПСП и постсинаптическое торможение по типу гиперполяризации. Тормозные влияния на сгибательные рефлексы оказывает преимущественно медиальная ретикулярная формация продолговатого мозга, а облегчающие – латеральные зоны РФ моста. Ретикулярная формация принимает участие в реализации многих функций организма. Так, РФ контролирует двигательную активность, постуральный тонус и фазные движения. В 1944 г. в США во время эпидемии полиомиелита (заболевания, нарушающего двигательную активность) основные структурные изменения были обнаружены в ретикулярной формации. Это навело американского ученого Г. Мэгуна на мысль об участии РФ в моторной активности. Основными ее структурами, отвечающими за этот вид деятельности, являются ядро Дейтерса продолговатого мозга и красное ядро среднего мозга. Ядро Дейтерса поддерживает тонус альфа- и гамма-мотонейронов спинного мозга, иннервирующих мышцы-разгибатели, и тормозит альфа- и гамма-мотонейроны мышц сгибателей. Красное ядро, напротив, тонизирует альфа- и гамма-мотонейроны мышц-сгибателей и тормозит альфа- и гамма-мотонейроны мышц-разгибателей. Красное ядро оказывает тормозное влияние на ядро Дейтерса, поддерживая равномерный тонус мышц-разгибателей. Повреждения или перерезка мозга между средним и продолговатым приводит к снятию тормозных влияний со стороны красного ядра на ядро Дейтерса, а значит и на тонус мышц разгибателей, который начинает преобладать над тонусом мышц-сгибателей, и возникает децеребрационная ригидность или повышенный тонус мышц, проявляющийся в сильном сопротивлении растяжению. Такое животное имеет характерную позу тела: запрокинутая голова, вытянуты передние и задние конечности. Поставленное на ноги, оно при малейшем толчке падает, так как отсутствует тонкая регуляция позы тела. 167 Ригидность исчезает, если разрушить ядро Дейтерса или произвести перерезку ниже ромбовидной ямки продолговатого мозга. Раздражение ретикулярной формации вызывает тремор, спастический тонус. РФ среднего мозга играет роль в координации сокращений глазных мышц. Получив информацию от верхних бугров четверохолмия, мозжечка, вестибулярных ядер, зрительных областей коры головного мозга, РФ ее интегрирует, что приводит к рефлекторным изменениям работы глазодвигательного аппарата, особенно при внезапном появлении движущихся объектов, изменении положения головы и глаз. Ретикулярная формация регулирует ряд вегетативных функций, в реализации которых принимают участие так называемые стартовые нейроны РФ, запускающие процесс возбуждения внутри определенной группы нейронов, отвечающих за дыхательные и сосудодвигательные функции. В РФ продолговатого мозга расположены два ядра, одно из них отвечает за вдох, другое – за выдох. Их деятельность контролируется пневмотаксическим центром РФ варолиева моста. Раздражением этих участков РФ можно воспроизвести различные дыхательные акты. Сосудодвигательный центр расположен в ромбовидной ямке дна IV желудочка, входящего в состав РФ. При электрораздражении определенных точек варолиева моста и продолговатого мозга возникают как прессорные, так и депрессорные сосудодвигательные реакции. Ретикулярная формация связана со всеми отделами коры мозга с помощью диффузной неспецифической проекционной афферентной системы, которая в отличие от специфической проводит возникшее на периферии возбуждение к коре больших полушарий медленно через последовательно связанные многонейронные системы. Раздражение РФ вызывает «реакцию пробуждения», а на электроэнцефалограмме – десинхронизацию альфа-ритма и переход его к более высокочастотному бета-ритму, а также ориентировочный рефлекс. Перерезка головного мозга ниже РФ вызывает картину бодрствования, выше – сна. РФ регулирует цикл «сон– бодрствование». Ретикулярная формация оказывает влияние на сенсорные системы мозга: на остроту слуха, зрения, обонятельные ощущения. РФ обладает высокой чувствительностью к таким веществам, как адреналин и углекислый газ, а также к лекарственным препаратам: барбитуратам и аминазину. Так, повреждение РФ и барбитуровый наркоз приводят к усилению сенсорных импульсов, которые в норме находятся под тормозным, регулирующим влиянием РФ. Восприятие различных ощущений при сосредоточении внимания на каком-либо другом ощущении, привыкание к повторяющимся раздражителям также объясняется ретикулярными влияниями. В ретикулярной формации продолговатого, среднего мозга и таламуса имеются нейроны, реагирующие на болевые раздражения от мышц и внутренних органов, при этом создается ощущение тупой боли. Вестибулярные ядра Все вестибулярные ядра функционируют вместе с ретикулярными ядрами моста для управления антигравитационными мышцами. Вестибулярные ядра посылают сильные возбуждающие сигналы к антигравитационным мышцам по латеральному и медиальному вестибулоспинальным трактам, расположенным в переднем столбе спинного мозга. Без этой поддержки вестибулярных ядер мостовая ретикулярная система потеряла бы большую часть своего возбуждающего влияния на осевые антигравитационные мышцы. Специфическая роль вестибулярных ядер, однако, состоит в избирательной регуляции возбуждения различных антигравитационных мышцах для подержания равновесия в ответ на сигналы от вестибулярного аппарата. У децеребрированного животного развивается спастическая ригидность. Когда ствол мозга животного пересекают ниже уровня середины среднего мозга так, чтобы интактными остались ретикулярные системы моста и продолговатого мозга, а также вестибулярная система, развивается состояние, называемое децеребрационной ригидностью. При этом ригидность развивается не во всех мышцах тела, а в антигравитационных мышцах шеи и туловища и в разгибателях ног. Причиной децеребрационной ригидности является блокада сильного влияния коры большого мозга, красных ядер и базальных ганглиев на ретикулярные ядра продолговатого мозга. Без этого влияния тормозное действие ретикулярной формации продолговатого мозга не проявляется, и мощная гиперактивность возбуждающей системы моста ведет к развитию ригидности. Мы увидим далее, что при других поражениях двигательных систем мозга, особенно системы базальных ганглиев, причины ригидности могут быть иными. Функции двигательных ядер среднего мозга. Децеребрационная ригидность. Ориентировочный рефлекс. Основными структурными образованиями среднего мозга являются: ядро блокового нерва – IV пара черепно-мозговых нервов (n. trochlearis), ядро глазодвигательного нерва (n. oculomotorius) – III пара черепно-мозговых нервов, красное ядро (n. ruber), черное вещество (substantia nigra), четверохолмие, ножки мозга и ядра ретикулярной формации. Блоковой нерв иннервирует верхнюю косую мышцу глаза, обеспечивает поворот глаза вниз-наружу. Глазодвигательный нерв отвечает за поднятие верхнего века, регуляцию движений глаза вверх, вниз, к носу и вниз к углу носа. Нейроны добавочного ядра глазодвигательного нерва (ядро Якубовича) регулируют просвет зрачка и кривизну хрусталика, обеспечивая процесс аккомодации. Красные ядра имеют связи с корой больших полушарий, подкорковыми ядрами, мозжечком, спинным мозгом. Они регулируют тонус мускулатуры Черное вещество располагается в ножках мозга, участвует в регуляции актов жевания, глотания и их последовательности, а также в координации мелких и точных движений пальцев рук, например, при письме, игре на скрипке, на рояле. Нейроны этого ядра синтезируют дофамин, поставляемый к базальным ядрам головного мозга (полосатому телу). Последний играет важную роль в контроле сложных двигательных актов. Поражение черного вещества приводит к дегенерации дофаминергических волокон, проецирующихся в полосатое тело, нарушению тонких движений пальцев рук, развитию мышечной ригидности и тремору (болезнь Паркинсона). Черное вещество оказывает тормозящее влияние на нейроны таламуса, посредником которого является медиатор ГАМК, и получает возбуждающие афферентные входы от субталамического ядра В каудальном отделе среднего мозга, но только у млекопитающих, находится так называемое «голубое пятно» (locus coeruleus). Основным медиатором этой структуры является норадреналин. Аксоны «голубого пятна» (ГП) связаны с корой больших полушарий, промежуточным мозгом, ядром ствола и моторными центрами спинного мозга. Аксоны ГП контактируют с альфа-мотонейронами передних рогов спинного мозга. Предполагают, что норадреналин является тормозным медиатором для альфа-мотонейронов. Во время фазы «быстрого сна» повышается активность нейронов ГП, которые с помощью норадреналина выключают мышечный тонус и фазические движения мускулатуры туловища и конечностей. Одностороннее разрушение ГП приводит к возникновению вращательных движений в сторону, противоположную разрушению. Вероятно, это результат нарушений связей между ГП и черным веществом. Помимо моторных, ГП выполняет гомеостатическую функцию за счет связей с чувствительными ядрами тройничного, языкогло- 136 точного и блуждающего нервов. ГП отвечает за постоянство внутренней среды организма. Нейроны ГП реагируют на изменения газового состава крови и ликвора, кроме того, они связаны с вегетативными центрами гипоталамуса. Верхние бугры четверохолмия – это первичный зрительный подкорковый центр, обеспечивающий зрительный ориентировочный рефлекс – поворот головы и глаз в сторону светового раздражителя, фиксацию взора и слежение за движущимися объектами. При повреждении или раздражении верхних бугров наблюдается нистагм глаз. Нижние бугры четверохолмия – это первичный слуховой подкорковый центр, участвующий в ориентировочном слуховом рефлексе – повороте головы в сторону источника звука. Двустороннее повреждение нижних бугров сопровождается потерей способности локализовать источник звука в пространстве. У человека рефлекс четверохолмия называется сторожевым, обеспечивает старт-реакцию на внезапные звуковые или зрительные раздражители. 1 2 |