А-Догель - Зоология беспозвоночных_2. Тип губки (spongia, или porifera)
 Скачать 14.77 Mb. Скачать 14.77 Mb.
|

|
348 стоногих (трилобиты, ракообразные), у многих ископаемых насекомых (см. рис. 362) и у некоторых современных форм (личинки термитов, некоторые богомолы, тараканы и т. д.; рис. 321). Переходом от ползающего к летающему послужил, быть может, лазающий образ жизни на деревьях, при котором насекомые, вероятно, часто делали прыжки с ветки на ветку, что способствовало дальнейшему развитию боковых выростов груди, которые вначале выполняли функции несущих плоскостей при парашютировании или планирующем полете. Дальнейшая дифференцировка и отчленение выростов от самого тела привели к выработке настоящих крыльев, обеспечивающих активный пропеллирующий полет. Брюшко — последний отдел тела насекомых (см. рис. 307). Количество сегментов, входящих в его состав, варьирует у разных представителей класса. Здесь, как и в других группах членистоногих, выявляется четкая закономерность: чем ниже в эволюционном отношении стоят те или иные представители, тем более полным набором сегментов они обладают. И действительно, максимальное число брюшных сегментов мы находим у самых низших скрыточелюстных (отр. Protura), брюшко которых состоит из 11 сегментов и заканчивается отчетливо различимым тельсоном (см. рис. 363, А, Б).У всех остальных насекомых часть сегментов редуцируется (обычно один или несколько последних, а иногда и самый первый), так что общее число сегментов может сокращаться до 10, а у высших форм (некоторые перепончатокрылые и двукрылые) до 4-5. 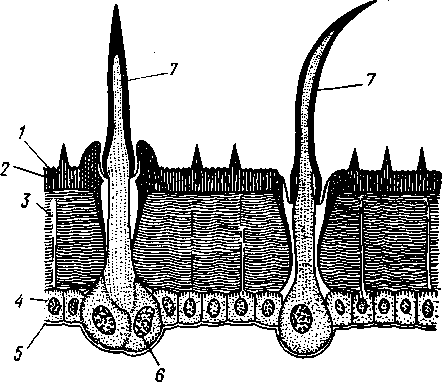 Брюшко обычно лишено конечностей. Однако вследствие происхождения насекомых от форм, обладавших ножками на протяжении всего гомономно расчлененного тела, нередко сохраняются на брюшке рудименты конечностей или же конечности, изменившие свою первоначальную функцию. Так, у отряда Protura, низших представителей бескрылых насекомых, имеются маленькие конечности на трех передних члениках брюшка. Сохраняются рудименты брюшных конечностей и у открыто-челюстных. У тизанур на всех сегментах брюшка имеются особые придатки— грифельки, на которых, как на полозьях, при движении насекомого брюшко скользит по субстрату (см. рис. 366, А). Одна пара . грифель- 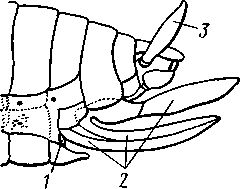 Рис. 322. Схема строения яйцеклада насекомого (из Бей-Биенко): / — половое отверстие. 2 — створки яйцеклада, 3 — цер- кн Рис. 323. Строение покровов насекомого (из Бей-Биенко) : /— наружный слой кутикулы, 2 — средний слой кутикулы, 3 —внутренний слой кутикулы, 4 — гиподерма, 5 — базальная мембрана. 6 — клетка, образующая волосок, 7 — волосок 349 ков на заднем конце тела сохраняется и у тараканов. Очень широко распространены, особенно у более примитивных форм (тараканы, саранчовые и т. п.), церки — парные придатки последнего сегмента брюшка, также являющиеся видоизмененными конечностями. По-видимому, сходное происхождение имеют и яйцеклады (рис. 322), встречающиеся у многих насекомых и состоящие из трех пар вытянутых створок. Покровы насекомых, как и всех остальных членистоногих, состоят из трех основных элементов — кутикулы, гиподермы и базальной мембраны (рис. 323). Кутикула выделяется клетками гиподермы, которая у скры-точелюстных насекомых часто превращается в синцитий. Кутикула насекомых трехслойна. В отличие от таковой ракообразных (с. 296) она несет наружный слой, содержащий липопротеинозые комплексы и препятствующий испарению воды из организма. Насекомые — сухопутные животные. Интересно отметить, что у водных и почвенных форм, обитающих в атмосфере, насыщенной парами воды, наружный слой или не выражен совсем, или развит очень слабо. Механическую прочность кутикуле придают белки, задубленные фенолами. Они инкрустируют средний, основной слой. На поверхности кутикулы находятся разнообразные, подвижно сочлененные с поверхностью тела выросты — тонкие волоски, чешуйки, щетинки. Каждое такое образование обыкновенно есть продукт выделения одной крупной гиподермальной клетки. Многообразие форм и функций волосков необычайно, они могут быть чувствительными, покровными, ядовитыми. Окраска насекомых в большинстве случаев зависит от присутствия в гиподерме или в кутикуле особых красящих веществ — пигментов. Металлический блеск многих насекомых относится к числу так называемых структурных окрасок и имеет другую природу. Особенности строения кутикулы обусловливают появление ряда оптических эффектов, которые основаны па сложном преломлении и отражении световых лучей. Покровы насекомых имеют разнообразного значения железы; они бывают одноклеточными и многоклеточными. Таковы вонючие железы (на груди клопов), защитные железы (у многих гусениц) и т. д. Наиболее часто встречаются линочные железы. Их секрет, выделяемый во время линьки, растворяет внутренний слой старой кутикулы, не затрагивая вновь образующихся кутикулярных слоев. Особыми восковыми железами у пчел, червецов и некоторых других насекомых выделяется воск. Мышечная система насекомых отличается большой сложностью и высокой степенью дифференциации и специализации отдельных ее элементов. Количество отдельных мышечных пучков часто достигает 1,5 — 2 тыс. Скелетные мышцы, обеспечивающие подвижность организма и отдельных его частей по отношению друг к другу, как правило, прикрепляются к внутренним поверхностям кутикулярных склеритов (тсргитам, стернитам, стенкам конечностей). По гистологическому строению почти все мышцы насекомых поперечнополосатые. Мышцы насекомых (в первую очередь это относится к крыловым мышцам высших групп 'насекомых: перепончатокрылым, двукрылым и т. п.) способны к необычайной частоте сокращений — до 1000 раз в секунду. Это связано с явлением умножения ответа на раздражение, когда на один нервный импульс мышца отвечает несколькими сокращениями. Богато разветвленная сеть трахейной дыхательной системы снабжает кислородом каждый мышечный пучок, что наряду с заметным повышением температуры тела насекомых при полете (за счет тепловой энергий, выделяемой работающими мышцами) обеспечивает высокую интенсивность обменных процессов, протекающих в мышечных клетках. 350 1 Пищеварительная система начинается небольшой ротовой полостью, стенки которой образуются верхней губой и совокупностью ротовых конечностей. У форм, питающихся жидкой пищей, она, по существу, заменена каналами (см. рис. 312,5), образующимися в хоботке и служащими для засасывания пищи и проведения слюны — секрета специальных слюнных желез. Стенки верхней части ротовой полости и следующей за ней трубчатой глотки соединяются со стенками головной капсулы с помощью мощных мышечных пучков. Совокупность этих пучков образует своеобразный мышечный насос, обеспечивающий продвижение пищи в пищеварительныю систему.  В заднюю часть ротовой полости, как правило, около основания нижней губы (максиллы II) открываются протоки одной или нескольких (до 3) пар слюнных желез. Содержащиеся в слюне ферменты обеспечивают начальные этапы процессов пищеварения. У кровососущих насекомых (муха цеце, некоторые виды комаров и т. п.) слюна часто содержит вещества, препятствующие свертыванию крови — антикоагулянты. В некоторых случаях слюнные железы резко меняют свою функцию. У гусениц бабочек, например, они превращаются в прядильные, которые вместо слюны выделяют шелковистую нить, служащую для изготовления кокона или для других целей. Пищеварительный канал насекомых (рис. 324), начинающийся глоткой, состоит из трех отделов: передней, средней и задней кишок. Передняя кишка может быть дифференцирована на несколько отличающихся по функциям и строению частей. Глотка переходит в пищевод, имеющий вид узкой и длинной трубки. Задний конец пищевода часто расширяется в зоб, особенно развитый у насекомых, питающихся жидкой пищей. У некоторых хищных жуков, прямокрылых, тараканов и т. д. за зобом помещается еще одно небольшое расширение передней кишки — жевательный желудок. Кутикула, выстилающая всю переднюю кишку, в жевательном желудке образует многочисленные твердые выросты в виде бугров, зубцов и т. п., способствующих дополнительному перетиранию пищи. Далее следует средняя кишка, в которой происходит переваривание и всасывание пищи; она имеет вид цилиндрической трубки. При начале средней кишки в нее нередко впадает несколько слепых выпячиваний Рис- 324- Пищеварительная система черного таракана (по Веберу): кишечника, или пилорических при- ;_м'юнные ж£иы. 2ЛищеволУ_аов. ДЗТКОВ, СЛуЖаЩИХ В ОСНОВНОМ ДЛЯ 4 — пилорические придатки, 5 — средняя о кишка, 6—мальпигисвы сосуды, 7 — зад- увеЛИЧеНИЯ ВСаСЫВаЮЩеИ ПОВерХНО- НЯЯ кишка, «-прямая кишка 351 сти кишечника. Стенки средней кишки часто образуют складки, или крипты. Обыкновенно эпителием средней кишки выделяется вокруг содержимого кишки непрерывная тонкая оболочка, так называемая пери-трофическая мембрана. В средней кишке происходит окончательное переваривание и усвоение пищевых веществ. Набор пищеварительных ферментов, характерный для того или иного вида насекомых, в первую очередь зависит от пищевого режима. Кровососы, пища которых в основном состоит из белков, обладают преимущественно протеолитическими ферментами, у бабочек, питающихся нектаром, имеются ферменты, воздействующие на сахара, и т. д. У многих растительноядных форм в кишечнике поселяются сим-биотические микроорганизмы (простейшие, бактерии и др.), обеспечивающие гидролиз клетчатки. Большую роль в процессах переваривания и всасывания пищевых веществ играет, по-видимому, перитрофическая мембрана, обеспечивающая транспорт ферментов внутрь пищевой массы, а продуктов пищеварения— в полость кишечника. Кроме того, она предохраняет эпителий средней кишки от механических повреждений. Средняя кишка переходит в эктодермальную заднюю, которая нередко отличается значительной длиной и подразделением на несколько участков. Здесь у большинства насекомых развиваются так называемые ректальные железы. О функциональных особенностях этих образований, как и всей задней кишки в целом, будет сказано ниже. Пища насекомых чрезвычайно разнообразна и охватывает почти все вещества растительного и животного происхождения. Среди насекомых имеются всеядные, например тараканы, которые питаются различными растительными и животными продуктами. Очень большое количество насекомых— чистые вегетарианцы и питаются различными частями растений от корня до плода включительно. Не менее многочисленны насекомые-хищники, поедающие других насекомых, червей, моллюсков и т. д. Наконец, имеются насекомые, питающиеся падалью и разного рода отбросами и продуктами гниения: навозом, гниющими растительными остатками и т. д. Некоторые из насекомых специализировались, казалось бы, на крайне малопитательной пище: перья, рог, воск и т. д. Органы выделения. Функции органов выделения у насекомых выполняются рядом образований. В первую очередь, это мальпигиевы сосуды, отходящие от пищевого канала на границе между средней и задней кишками (см. рис. 324). Мальпигиевы сосуды Tracheata развиваются за счет задней кишки и имеют соответственно эктодермальное происхождение. Их полость выстлана низким однослойным эпителием. Количество мальпигиевых сосудов колеблется от 2 до 200 и более или менее постоянно у представителей разных систематических групп. Иногда мальпигиевы сосуды обладают некоторой подвижностью, что обеспечивает постоянную смену омывающей их гемолимфы. Из гемолимфы продукты обмена в виде растворенных в воде солей мочевой кислоты поступают в полость сосудов, где образуется труднорастворимая мочевая кислота, кристаллы которой сразу же начинают выпадать в осадок. Освобождающаяся при этом вода всасывается стенками сосудов и поступает обратно в гемолифму. Особенно интенсивно эти процессы идут в задней кишке, куда поступают продукты обмена из мальпигиевых сосудов. Упоминавшиеся выше ректальные железы являются основным местом всасывания воды. Почти сухие кристаллы мочевой кислоты вместе с непереваренными остатками пищи выводятся наружу через анальное отверстие. 352 1 Описанный выше процесс выведения продуктов обмена в виде сухих кристаллов, а не в растворе, позволяет насекомым очень экономно расходовать воду, поступающую в организм. Последнее особенно важно для видов, которые существуют в условиях сильного дефицита влаги. У форм, обитающих в среде с повышенной влажностью или поглощающих большое количество жидкой пищи (например, тли, сосущие соки растений), реабсорбция воды не наблюдается. Кроме мальпигиевых сосудов выделительную роль играет жировое тело, развитое у насекомых сильнее, чем у многоножек. Это рыхлая ткань, обильно пронизанная трахеями . Часть составляющих ее клеток выполняет экскреторную функцию — в них откладывается мочевая кислота в виде круглых конкреций. Жировое тело относится к числу «почек накопления», в которых продукты обмена постепенно накапливаются, а не выводятся во внешнюю среду. Не следует, однако, забывать, что главная функция жирового тела — отложение запасных питательных веществ. Благодаря образованию таких запасов насекомые могут подолгу голодать (например, клопы — до 6 месяцев и больше). Сложные процессы метаморфоза насекомых, особенно на стадии куколки, когда организм не питается, также осуществляются за счет энергетических веществ, накопленных в жировом теле. Говоря об органах выделения, следует упомянуть о перикардиальных клетках, или нефроцитах, которые часто наблюдаются у насекомых. Это парные более или менее метамерно расположенные по бокам сердца группы клеток, способные поглощать из полости тела введенные туда посторонние вещества, например кармин. Помимо перикардиальных клеток у некоторых низших насекомых (тизануры, прямокрылые) найдены расположенные под сердцем в виде особых скоплений амебоидных клеток фагоцитарные органы. Последние способны захватывать различные мелкие твердые частицы, например зерна туши, введенной в полость тела. Видоизмененными участками жирового тела являются органы свечения некоторых насекомых, например жуков-светлячков Lampyris (Иванов червячок; рис. 325). Эти участки жирового тела залегают под прозрачным кутикулярным покровом брюшка; свечение их зависит от присутствия в клетках особого вещества люциферина. При наличии кислорода под воздействием особого фермента люциферазы происходит  Рис. 325. Светляк, или Иванов червячок, Lampyris noctiluca. А — самец; Б личинка (из Брема) 12-763 353 окисление люциферина, причем химическая реакция сопровождается испусканием света. Процессы свечения насекомых находятся под контролем нервной системы. Нервная система насекомых (рис. 326), как и у прочих членистоногих, исходно построена по типу брюшной нервной цепочки, однако может достигать очень высокого уровня развития и специализации. Центральная нервная система включает головной мозг, подглоточ-ный ганглий и сегментарные ганглии брюшной нервной цепочки, расположенные в туловище. Головной мозг состоит из трех участков: передний— протоцеребрум, средний — дейтоцеребрум и задний — тритоце-ребрум. Протоцеребрум и дейтоцеребрум иннервируют соответственно глаза и сяжки насекомых, т. е. придатки акрона. Тритоцеребрум принадлежит вставочному, или интеркалярно-  2 А .1 - му, сегменту, который отвечает сегмен ту второй пары антенн раков. Голов ной мозг отличается очень сложным гистологическим строением, так как каждый его отдел, в свою очередь, слагается из нескольких ганглиозных скоплений, разделенных прослойками из нервных волокон. Особенно важными ассоциативными центрами головного мозга считаются «грибовидные тела», располагающиеся в протоцеребруме. Впрочем, сложность структуры характеризует мозг не всех насекомых, а главным образом тех, жизнь которых отличается сложностью и разнообразием жизненных функций. Поэтому мозг развит сложнее всего у общественных насекомых: муравьев, пчел, термитов. Эта закономерность у них прослеживается даже в пределах одного вида, представленного несколькими «кастами», отличающимися по сложности жизненных отправлений. У рабочих муравьев, например, грибовидные тела развиты значительно сильнее, чем у цариц и самцов (рис. 327). Рис. 326. Схема строения нервной системы насекомого (из Вюрмбах): 1 — протоцеребрум, 2 — нейросекретор-ные клетки, 3 — оптическая область мозга, 4 — дейтоцеребрум, 5 — антен-нальный нерв, 6 — тритоцеребрум, 7 — кардиальные тела, 8 — прилежащие тела, 9 — окологлоточные коннективы, 10—подглоточный ганглий, II — нервы, идущие к ротовым конечностям, 12 —ганглии грудных сегментов, 13 — ганглии брюшных сегментов, 14 — непарный нерв симпатической системы Брюшная нервная цепочка состоит из сложного подглоточного ганглия, посылающего нервы к трем парам ротовых конечностей, из трех крупных обособленных грудных ганглиев и брюшных ганглиев, количество которых может варьировать. Наиболее полное их число — 11 — наблюдается только на самых ранних стадиях эмбриогенеза некоторых насекомых — тараканы, медведки, жуки и т. д. У большинства насекомых ганглии брюшной цепочки концентрируются в продольном направлении (рис. 328), так что во взрослом состоянии даже у 354 1 самых примитивных форм не встречается более 8 брюшных ганглиев. Причем последний, -VIII ганглий сохраняет следы своего сложного происхождения за счет слияния нескольких ганглиев. Однако у многих насекомых процесс концентрации ганглиев заходит значительно дальше (рис. 328). Возникают сложные брюшные и грудные ганглиозные массы. В ряде случаев все ганглии грудного и брюшного отделов могут сливаться, образуя массу, расположенную в груди, тогда как в брюш- 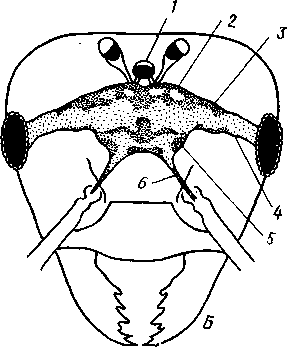 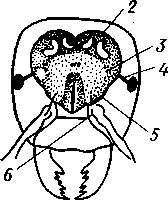 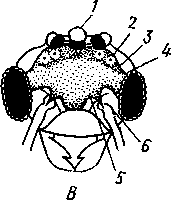 Рис. 327. Головной мозг муравья Lasius brevicornis. A — мозг рабочей особи; Б — мозг самки; В — мозг самца (по Уилеру): / — срединный лобный глазок, 2 — грибовидные тела мозга, наиболее развиты у рабочих пчел И рудиментарны у самцов, 3 — зрительные доли, 4 — зрительный нерв, 5 — обонятельные доли, 6 — сяжковый нерв. Все рисунки сделаны при одинаковом увеличении 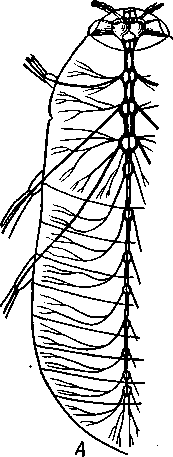 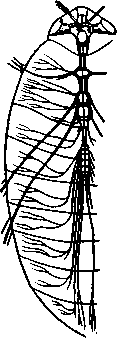 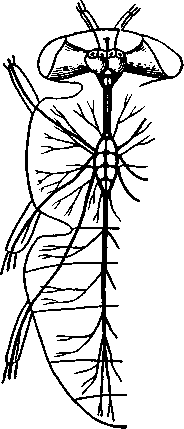 •Б В Рис. 328. Нервная система насекомых. А — жука Lygistopterus san-guineus; Б — жука Gyrinus notator; В — мухи Sarcophaga carnaria (по Брандту) 355 12» 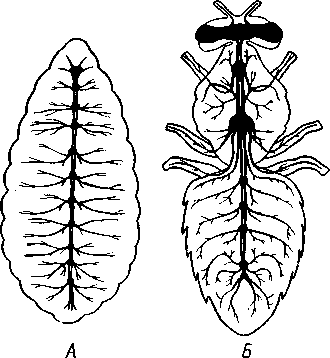 ке остаются только нервы. Обычно нервная цепочка личинок богаче расчленена, чем таковая взрослых насекомых: взрослая пчела имеет всего 6 ганглиев вместо личиночных 10 (рис. 329). Насекомые обладают системой отходящих от головного мозга симпатических нервов, которые регулируют работу внутренних органов и мышечной системы. Практически во всех отделах центральной нервной системы (головной мозг, подглоточный ганглий, брюшная нервная цепочка) имеются нейросекреторные клетки. Синтезируемый в них нейросекрет транспортируется по аксонам в особые образования — прилежащие и карди-альные тела, а затем поступает в гемолимфу. Кардиальные и прилежащие тела располагаются над кишечником сразу же за головным мозгом. Они выполняют функции желез внутренней секреции. Рис. 329. Развитие брюшной нервной цепочки у домашней пчелы Apis mel-lifera. A — личинка; Б — взрослое насекомое (по Бланшару и Сподграссу) Нейросекреты играют важную роль в гормональной системе насекомых: они регулируют деятельность всех остальных эндокринных органов, гормоны которых обеспечивают нормальное осуществление развития организма, течение обменных процессов, линьки и т. д. (с. 374). Органы чувств насекомых достигают большой сложности и многообразия. Это в первую очередь определяется как общим высоким уровнем организации насекомых, так и их сложным поведением, для осуществления которого организм должен получать исчерпывающую и точную информацию об окружающем мире. Насекомые способны воспринимать самые различные раздражения — механические, звуковые, химические, зрительные и т. д. Для этого у них имеются специализированные рецепторы. Морфологической и функциональной основой органов чувств являются сенсиллы, разбросанные по телу насекомых поодиночке или собранные в более или менее крупные скопления (органы слуха, глаза и т. п.). Все сенсиллы возникли из одного исходного типа путем специализации для восприятия того или иного раздражения. Каждая сенсилла состоит из нескольких элементов. Это одна или несколько чувствительных (рецепторных) клеток. От каждой клетки берет начало центральный отросток, идущий к нервной системе, и периферический отросток. На конце последнего в большинстве случаев располагается видоизмененный жгутик, у которого отсутствуют две центральные фибриллы. Наружные части сенсиллы представлены кутикулярным образованием, строение которого зависит от выполняемых им функций. Жгутик чувствительной клетки направлен к кутикулярной части сенсиллы. Здесь его конец входит внутрь особого канала или трубочки, которые фиксируют положение жгута. Наиболее простыми являются, по-видимому, сенсиллы, воспринимающие механическое раздражение (рис. 330). Основу такой сенсиллы составляет волосок, подвижно сочлененный с кутикулой. Изменение положения волоска при соприкосновении с твердыми предметами или под 356 давлением воды или воздуха сразу же передается жгутику и чувствительной клетке, в которой возникает возбуждение. Такие осязательные или тактильные сенсиллы разбросаны по всей поверхности тела насекомого. По существу, механорецепторными являются и так называемые хор-дотональные сенсиллы, или сколпофоры(рис. 331), служащие для вос- 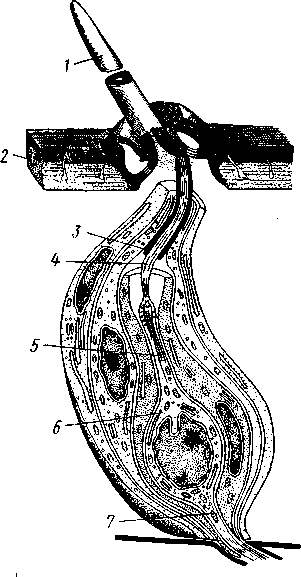 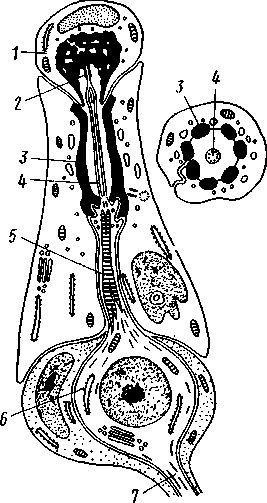 Рис. 330. Строение механорецеп-торной сенсиллы насекомых (рис. В. Иванова): / — чувствительный волосок, 2 — кутикула, :3 — кутикулярная трубочка, фиксирующая жгутик, 4 — видоизмененный жгутик, 5—периферический отросток чувствительной клетки, 6 — чувствительная клетка, 7 — центральный отросток чувствительной клетки Рис. 331. Схема строения хордо-тоналыгой сенсиллы насекомых (по В. Иванову): / — шапочковая клетка, 2 — кутикулярная трубочка, фиксирующая жгутик, 3 —сколопоидное тельце, 4 — видоизмененный жгутик, 5 — периферический отросток чувствительной клетки, 6 — чувствительная клетка, 7 — центральный отросток чувствительной клетки приятия колебаний различной частоты. Они в первую очередь входят в состав органов слуха насекомых. Хордотональные сенсиллы отличаются некоторыми особенностями строения. Вокруг периферического отростка чувствительной клетки и жгутика, заключенного в кутикулярную трубочку, формируется плотный фибриллярный чехол — сколопоидное тельце, имеющее вид блестящего штифтика. Сверху сенсиллу прикрывает особая шапочковая клетка, от нижней кутикулизированной поверхности которой и берет начало трубочка, фиксирующая жгут. Хордотональные сенсиллы располагаются поодиночке или группами, образуя хордото-нальные органы. Чаще всего они примыкают к внутренней поверхности мягких участков кутикулы (в сочленениях конечностей или же между сегментами тела). Иногда они погружены в глубь тела и расположены 35Z -на тонких мембранных трубках, натянутых между двумя отдельными участками кутикулы (рис. 332). 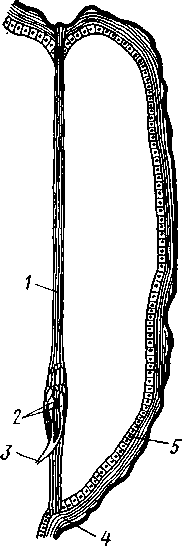 Функции хордотональных органов, по-видимому, различны. В тех случаях, когда сенсиллы примыкают к кутикуле, они, как правило, служат для восприятия низкочастотных вибраций. Правда, в отдельных случаях (хордотональные органы, расположенные в антеннах комаров) они чувствительны и к колебаниям высокой частоты. Внутренние хордотональные органы, вероятно, регистрируют изменения давления и механических напряжений, возникающих в теле насекомого. Настоящими слуховыми органами насекомых являются тимпанальные органы, в которых сколпо-форы связаны с тонкими кутикулярными мембранами (тимпанальными мембранами), играющими роль барабанных перепонок. |