А-Догель - Зоология беспозвоночных_2. Тип губки (spongia, или porifera)
 Скачать 14.77 Mb. Скачать 14.77 Mb.
|

|
Экология. Все плеченогие — обитатели морей. Они широко распространены и встречаются на различных глубинах. Питаются Brachiopoda различными мелкими животными и органическими остатками, взвешенными в воде. Большинство совершенно неподвижно прикреплено к суб- 516 страху: Lingula (из Ecardines) имеет длинный мясистый свободный стебелек, погружающийся в мягкий грунт. Плеченогих следует считать вымирающей группой. Как уже сказано, число известных науке вымерших видов превышает 10000. Раковины Brachiopoda появляются с до-кембрийских отложений. Попадаясь в большом количестве, начиная с самых древних эпох, плеченогие имеют очень большое значение в палеонтологии, ибо служат «руководящими окаменелостями» для многих пластов. Интересно, что некоторые роды плеченогих обнаруживают в течение огромных промежутков времени крайне малую изменчивость и большую стойкость организации. Так, Lingula, встречающаяся в настоящее время очень часто у берегов Японии, сохранила все родовые признаки с силурийского периода. Среди ископаемых плеченогих особенно любопытны виды, жившие в сильно подвижной воде и прикреплявшиеся брюшной створкой. Последняя принимает вид высокого толстостенного конуса, в котором помещается животное, тогда как спинная створка низводится до степени небольшой крышечки, замыкающей вход в конус (Richthofenia). Классификация. Класс Brachiopoda делится на два подкласса. ПОДКЛАСС I. БЕЗЗАМКОВЫЕ (ECARDINES, ИЛИ INARTICULATA) Наиболее древние плеченогие, известные еще из докембрийских отложений. Замковое соединение створок отсутствует. Стебелек иногда преобразуется в сократимую ногу, служащую для рытья. Кишечник изогнут и открывается наружу анальным отверстием. Выходящая в воду личинка заключена в двустворчатую раковину. Представители: Lingula, ведущая роющий образ жизни, и др. ПОДКЛАСС II. ЗАМКОВЫЕ (TESTICARDINES, ИЛИ ARTICLJLATA) Филогенетически более молодые плеченогие, появляющиеся в нижнем кембрии. Створки раковины соединяются с помощью замка. Пищеварительная система слепо замкнута, анальное отверстие отсутствует. Раковина появляется только в процессе метаморфоза личинки после прикрепления последней к субстрату. Представители: Rhynchonella, обитающая в Белом море; Magellania (см. рис. 504, 505); ископаемый Spirifer (см. рис. 507) и др. КЛАСС III. ФОРОНИДЫ (PHORONIDEA) Очень небольшая, состоящая всего из 18 видов группа морских животных, имеющих вытянутую, колбасообразную форму и прячущихся внутри выделяемой ими трубки, из которой высовывается только передний конец тела (рис. 509), снабженный, как и у мшанок, подковообразным лофофором с венчиком мерцательных щупалец (рис. 510), подгоняющих пищу ко рту. Последний прикрыт эпистомом. Немного позади рта лежит порошица, так что кишечник образует петлю. Дыхание совершается преимущественно через щупальца, выделение — посредством пары трубчатых органов, снабженных ресничной воронкой. Предполагается, что по своей природе это нефромиксии (см. с. 258). Через эти же органы наружу выводятся и половые продукты. Половая система гермафродитна и состоит из яичника и семенника, лежащих у заднего конца тела. Нервная система состоит из околоротового кольца и продольного ствола, проходящего асимметрично по левой 517 стороне тела. Органы чувств представлены особым лофофоральным органом, имеющим вид язычка и расположенным позади эпистома. Полость тела вторичная и состоит из трех отделов: переднего — эпи-стомального, среднего — кольцевидного, посылающего каналы в лофо-фор, и заднего — большого, занимающего все тело. Кровеносная система состоит из околоротового кольца и двух продольных сосудов, переходящих один в другой на заднем конце тела. 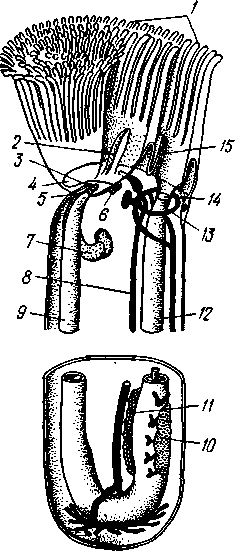 Рис. 509. Форонис — Phoronis psammopluta в своей трубке, состоящей из зерен песка (по Сели Лоншану) Рис. 510. Схема строения Phoronis (по Дсляжу и Эроуару): 1 — внутренний и наружный ряды щупалец лофофора, 2 — лофофораль-ный орган, выполняющий, вероятно, чувствительную функцию, 3 — выделительное отверстие, 4 — нервное кольцо, 5—анус, 6 — нервный ганглии, 7 — нефромиксии, £ — спинной кровеносный сосуд, 9 — средняя кишка, 10 — семенник, // — яичник. 12—пищевод, 13 — лофофоральный кровеносный сосуд, 14 — ротовое отверстие, 15 — эпистом Развитие Phoronis ведет к образованию плавающей личинки актинотро-хи, похожей некоторыми признаками организации на трохофору, и сопровождается метаморфозом. Туловище «червя» образуется за счет разрастания брюшной стороны тела личинки (рис. 511). Форониды обладают и бесполым размножением, которое осуществляется в виде поперечного деления. Образующиеся особи позднее достраивают недостающие участки тела. У нас представители Phoronidea найдены в Черном море и в морях Дальнего Востока. 518 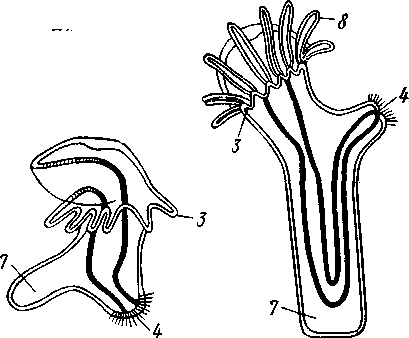  Рис. 511. Развитие Phoronis. A — личинка актинотроха сбоку; Б и В — стадии метаморфоза (по Маркусу): I — теменная пластинка, 2 — рот, 3 — личиночные щупальца, 4 — анус, 5 — кишка. 6 — передняя кишка, 7 — брюшной вырост тела — будущее туловище «червя», 8 — дефинитивные щупальца Дополнение к типу Tentaculata КЛАСС ВНУТРИПОРОШИЦЕВЫЕ (ENTOPROCTA, ИЛИ KAMPTOZOA) Небольшая (60 видов) группа сидячих, в основном морских, животных до недавнего времени относилась к классу Bryozoa и называлась внутрипорошицевыми мшанками. Современные авторы считают положение этой группы в системе менее определенным, поэтому мы провизорно помещаем ее в виде дополнения к типу Tentaculata. Entoprocta — мелкие животные; длина особи не превышает 1 мм. Тело чашевидное, один полюс его несет венчик мерцательных щупалец, окружающих рот и заднепроходное отверстие (в отличие от Bryozoa), другой продолжается в сократимый стебелек, снабженный продольным мускулом (рис. 512). Кишечник подковообразный, между ртом и порошицей находится ганглий, прилегающий к пищеводу (рис. 513). Имеется пара протонефридиев. Entoprocta или раздельнополы, или гермафродиты, причем половые железы имеют вид маленьких парных мешочков. Настоящего целома нет, промежутки между внутренними органами заняты паренхимой. Помимо полового размножения Entoprocta обладают и бесполым, а именно почкованием. Образующиеся на теле почки большей частью остаются в связи с материнской особью — возникают колонии Entoprocta (см. рис. 512). У представителей рода Loxosoma колоний нет; почки отделяются от матери и переходят к самостоятельной жизни. При половом размножении Entoprocta в результате спирального дробления получается личинка, снабженная венчиком ресниц и ресничным султаном и напоминающая многих личинок мшанок. С другой стороны, личинка похожа и на трохофору (кл. Polychaeta). Мезодерма личинки образуется из клетки 4d, дающей пару малоклеточных мезодермальных полосок, распадающихся на отдельные клетки, так что образования целома у Entoprocta не наблюдается. 519 Представители: Pedicellina, Loxosoma (моря), Urnatella (пресные воды Северной Америки). 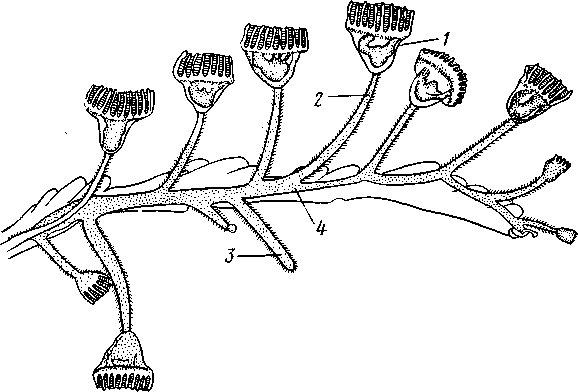 Рис. 512. Часть колонии Pedicellina cernua (из Абрикосова): / — тело зооида — чашечка, видны щупальца и кишечник, 2 — стебелек, 3 — стебелек сброшенной чашечки, 4 — столон колонии 10 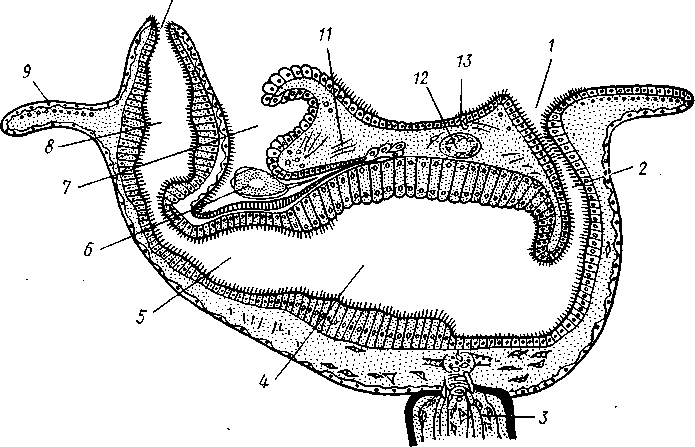 Рис. 513. Разрез чашечки Pedicellina cernua (из Абрикосова): / — рот, 2 — пищевод, 3 — стебелек, 4 — желудок, 5 — кишка, 6 — яйцо, выходящее из полового отверстия, 7—«выводковая камера», 8—прямая кишка, 9 — щупальце, 10 — анальное отверстие, Ч — паренхима, 12 — нервный узел, 13 — выделительное отверстие 520 Филогения типа Tentaculata Филогения щупальцевых далеко не выяснена. Наличие у них вторичной полости тела свидетельствует, что они выше всех низших червей и по высоте организации стоят на одной ступени с кольчецами. О том же свидетельствует и наличие у некоторых групп хорошо развитой кровеносной системы, иногда даже с центральным пропульсаторным аппаратом (кл. Brachiopoda). Взаимоотношения их с кольчецами не могут, однако, считаться особенно близкими ввиду того, что основные моменты их развития носят иной характер: отсутствует спиральное дробление, неясна тело-бластическая закладка мезодермы. С другой стороны, у многих щупальцевых имеются черты, сближающие их с другим большим разделом животного царства, а именно с вто-ричноротыми. Такова склонность к образованию разных зачатков не отщеплением плотных клеточных масс или отдельных клеток (телоблас-тов), а при помощи эпителиальных впячиваний и углублений. У мшанок ганглий и средняя кишка закладываются при почковании в виде впячиваний эктодермы, у некоторых плеченогих мезодерма отчетливо образуется за счет боковых выпячиваний средней кишки, т. е. совсем таким же способом,• как у вторичноротых (с. 555). К тому же разделение тела щупальцевых на три сегмента вполне напоминает сегментацию молодых стадий при развитии иглокожих, полухордовых и хордовых. Поэтому впредь до дальнейшего выяснения их филогении мы будем считать щупальцевых своеобразным типом целомических животных, сильно видоизмененным в результате сидячего образа жизни и расположенным у места расхождения двух главных ветвей билатеральных животных, т. е. Protostomia и Deuterostomia. В пределах самого типа щупальцевых более примитивными формами правильнее считать кл. Phoronidea, так как они в большей степени сохранили способность к движению: нет еще прирастания самого тела к субстрату. Мшанки же обнаруживают признаки вторичного упрощения и специализации (вторичная утрата энтодермы— с. 502; исчезновение органов выделения, кровеносной системы). Особенно отчетливо все это проявляется у представителей подкл. Gymnolaemata, тогда как покрыторотые мшанки (подкл. Phylactolaema-ta), сохранившие более сложное строение, по-видимому, стоят ближе к исходному типу. Класс Brachiopoda располагается обособленно от первых двух классов, хотя и обнаруживает некоторое родство с мшанками. Строение околоротового щупальцевого аппарата, наличие эпистома, характер и сегментация полости тела, сидячий образ жизни и некоторые особенности развития свидетельствуют в пользу сближения этих групп. Однако пле-ченогие обладают некоторыми признаками более сложной организации: кровеносная система, сердце, печень. Впрочем, некоторые зоологи отказывают плеченогим в родстве с мшанками и рассматривают класс Brachiopoda как самостоятельный тип, занимающий промежуточное положение между ветвями первичноротых и вторичноротых (Protostomia и Deuterostomia). Положение кл. Entoprocta в системе, как сказано, неопределенно. Одни авторы на основании сходства образа жизни, строения щупалец, сходства личинок и т. д. сближают его с мшанками. В таком случае следует считать, что отсутствие иелома у Entoprocta вторично, так же как и развитие в теле паренхимы. Другие авторы придают большое значение отсутствию целома и присутствию протонефридиев, относя кл. Entoprocta к низшим червям. 521 ТИП ИГЛОКОЖИЕ (ECHINODERMATA) Иглокожие — обширная, около 5000 видов, группа морских донных животных, большей частью свободноподвижных, реже прикрепленных ко дну посредством особого стебелька (рис. 514). Тип Echinociermata характеризуется следующими признаками. 1. Иглокожие обладают радиальной и притом обычно пятилучевой симметрией, однако их предки были билатерально симметричными животными. 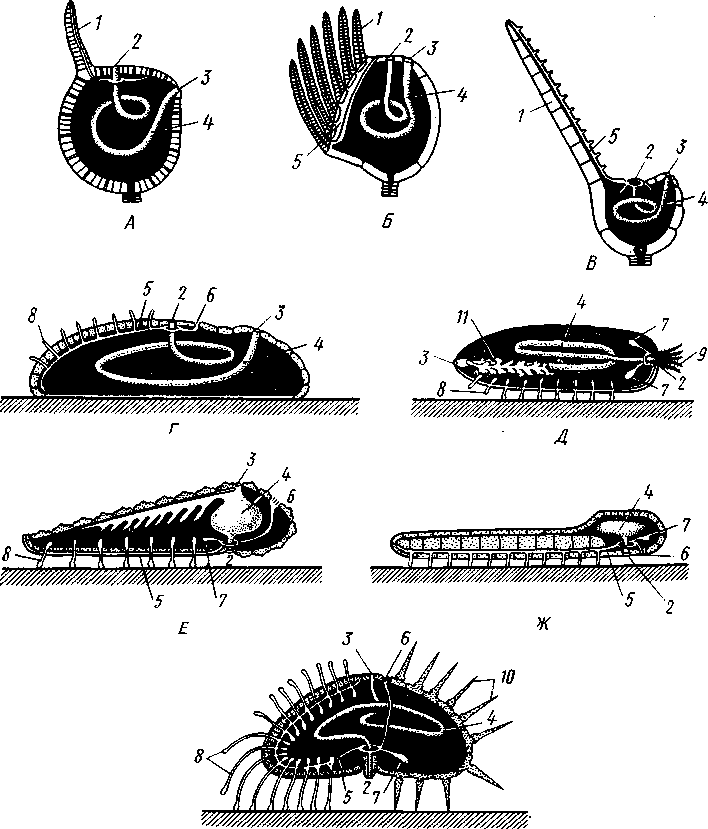 Рис. 514. Схема строения различных классов иглокожих. А — цистоидеи; Б — бластоидеи; В — морские лилии; Г — эдриоастероидеи; Д — голотурии; Е — морские звезды; Ж — офиуры; 3 — морские ежи: 7 — руки, 2 — рот, 3— анус, 4 — кишечник, 5 — амбулакральная система, 6 — мадрепоровая пластинка, 7 — полиевы пузыри, 8—амбулакральные ножки, 9 — щупальца, 70 — иглы. il — водное легкое 522 2. В подкожном соединительном слое иглокожих развивается скелет из известковых пластинок с торчащими на поверхности тела шипами, иглами и т. п. 3. Внутренние органы лежат в обширной полости тела (целоме). Одной из наиболее оригинальных черт строения иглокожих следует считать сложную дифференциацию части целома на ряд систем, в том числе образование за счет целома амбулакральной (воднососудистой) системы органов движения. 4. Имеется кровеносная система; органы дыхания слабо развиты или отсутствуют; специальных органов выделения нет. 5. Нервная система примитивна и частью залегает непосредственно в толще кожного эпителия или в эпителии участков стенки тела, впятившихся внутрь. 6. Иглокожие раздельнополы. Яйца испытывают полное радиальное дробление. В развитии иглокожих имеется характерная личинка диплев-рула, испытывающая сложный метаморфоз. Тип Echinodermata делится на несколько классов, группирующихся в два подтипа: Пельматозои (Pelmatozoa) и Элеутерозои (Eleutherozoa). К первому подтипу относятся ископаемые классы: карпоидеи (Сагро-idea), шаровики (Cystoidea), морские бутоны (Blastoidea), эдриоастеро-идеи (Edrioasteroidea) и современный класс — морские лилии (Cri-noidea). Ко второму подтипу — классы морские звезды (Asteroidea), сфиуры (Ophiuroidea), офиоцистии (Ophiocistia — ископаемая группа), морские ежи (Echinoidea) и голотурии (Holothuroidea). Мы рассмотрим преимущественно современные классы и по соображениям удобства начнем с Eleutherozoa, ПОДТИП ЭЛЕУТЕРОЗОИ (ELEUTHEROZOA) Высшие свободноподвижные иглокожие. Тело звездообразной, шаровидной или червеобразной формы. Скелет обычно в виде панциря, но может более или менее редуцироваться. Ротовое и анальное отверстия находятся на противоположных полюсах тела. Амбулакральная система обычно служит для движения. КЛАСС I. МОРСКИЕ ЗВЕЗДЫ (ASTEROIDEA) Морские звезды встречаются на разных глубинах; одни виды живут на глубинах в тысячи метров, другие у самых берегов, оставаясь иногда во время отлива по нескольку часов без воды. Звезды во многих отношениях выносливы, но (подобно другим иглокожим) крайне чувствительны к степени солености воды, нуждаясь в воде нормальной океанической солености (около 3%). Поэтому они отсутствуют в Черном и Балтийском морях. Звезды могут достигать больших размеров, до 70 см и более от конца одного луча до конца луча, ему противоположного; нередко ярко и пестро окрашены. Число видов — более 1700. Строение и физиология. Тело морских звезд имеет вид пятилучевой звезды (рис. 515), в которой различают центральный диск и пять лучей, или рук. Однако встречаются звезды, имеющие больше пяти лучей: с шестью (Hexaster) или же с девятью, одиннадцатью, тринадцатью лучами и более. Особенно большое число лучей (более 30) имеют звезды из семейства Brisingidae. Для удобства ориентировки в теле иглокожих различают, во-первых, линии, идущие от центра к концу лучей, называемые радиусами или 523 радиальными линиями; во-вторых, линии, заканчивающиеся на краю диска между соседними лучами, которые обозначаются как интеррадиусы или интеррадиальные линии. Тело звезды сплющено по направлению оси симметрии. В центре одной из плоских сторон помещается рот (оральная сторона), в центре другой — порошица (аборальная сторона). Животное ползает по дну ртом книзу. Ползание совершается при помощи особых отростков, ам- 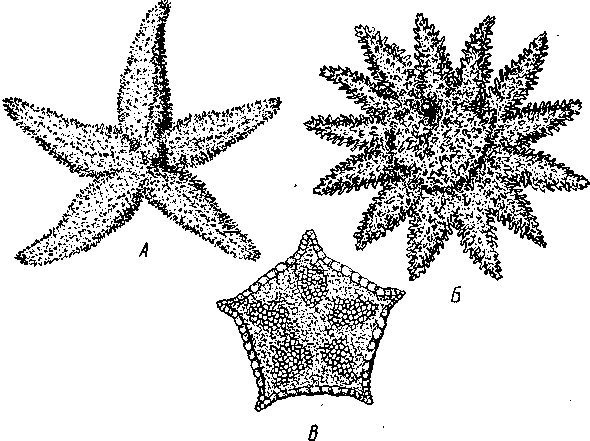 Рис. 515. Морские звезды. А—Asterias rubens; Б — Crossaster рар-posus; В — Ceratomaster granularis (из Аверинцева) булакральных ножек, расположенных на дне амбулакральной борозды на нижней (оральной) стороне каждого луча. Стенка тела состоит из однослойного обычно ресничного эпителия и слоя подстилающей его соединительной ткани; под соединительной тканью залегает перитонеальныи эпителий, ограничивающий вторичную полость тела, или целом, в котором помещаются все внутренности. Целом иглокожих состоит из нескольких самостоятельно закладывающихся у личинки участков, которые испытывают различные изменения и служат для образования нескольких систем полостей. В подкожной соединительной ткани развивается известковый скелет сначала в виде микроскопических телец, которые позднее сливаются в более крупные и правильно расположенные пластинки. Скелет сильнее развит на ротовой, оральной, стороне тела. В каждом луче имеются два ряда амбулакральных пластинок, которые соединены между собой попарно и прикрывают, наподобие двускатной кровли, амбулакральную борозду ротовой (оральной) стороны. Соседние пары амбулакральных пластинок соединены подвижно при помощи мышц. Кнаружи от амбулакральных с каждой стороны луча имеется по ряду адамбулакральных пластинок, а над последними, на боковой стороне луча, по 1—2 ряда краевых, или маргинальных, пластинок (рис. 516, 517). Скелет аборальной стороны представлен большей частью лишь многочисленными узкими известковыми перекладинами. Среди них в одном |