А-Догель - Зоология беспозвоночных_2. Тип губки (spongia, или porifera)
 Скачать 14.77 Mb. Скачать 14.77 Mb.
|

|
частью обладают небольшими размерами (несколько сантиметров), хотя среди них имеются виды, достигающие 2,5 м длины. Строение и физиология. Тело длинное, червеобразное, делится на три участка: хоботок, воротничок и туловище (рис. 555). Хоботок имеет характерную форму желудя или более или менее вытянут; его узкое основание, или шейка, охватывается следующим отделом тела — воротничком. Короткий воротничок отделен легкой перетяжкой от туловища, составляющего большую часть длины всего тела. Передняя часть туловища про- резана по бокам двумя рядами узких поперечных жаберных щелей. Все тело кишечнодышащих одето нежным однослойным мерцательным кожным эпителием Рис. 555. Строение кишечнодышащих Enteropneusta (по Шпенгелю). А — Saccoglossus kowalevskyi, видно разделение тела на хоботок, воротничок и туловище; Б — продольный разрез переднего конца Ptychodera minuta: 1 — жаберные щели, 2 — сплетение кровеносных сосудов, 3 — перикардий, 4 — хоботная пора, 5 — спинной нервный тяж, 6 — спинной сосуд, 7 — жаберная кишка, 8 — брюшной сосуд, 9—пищевод, 10 — глотка. // — сердечная лакуна. 12 — нотохорд, 13—продольные мускулы хоботка 562 (рис. 556), клетки которого покоятся своими основаниями на тонкой бесструктурной базальной перепонке (мембране). Кнутри от мембраны располагается мускулатура, состоящая из наружного слоя кольцевых и внутреннего — продольных гладких мышечных волокон. 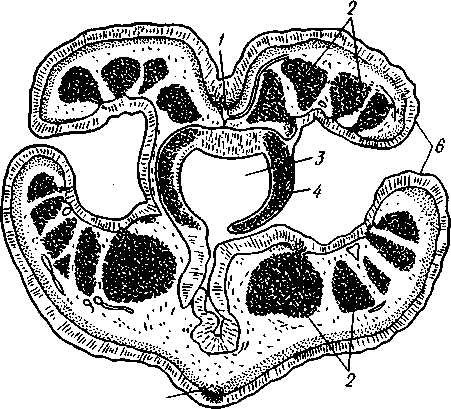 Пищеварительная и дыхательная системы. Рот находится на брюшной стороне тела у основания хоботка, на границе последнего с воротничком (см. рис. 555). Рот ведет в короткую глотку, лежащую на протяжении воротничка. От спинной стороны самого начала глотки отходит кпереди, внутрь хоботка, слепое выпячивание кишечника, называемое нотохордом. Он имеет узкий просвет, стенки его состоят из одного слоя очень крупных вакуолизированных клеток. Сравнение нотохорда с хордой позвоночных про водится на основании сходства их положения (над кишкой), строения (состоят из крупных ва- $ Рис. 556. Поперечный разрез через жаберный отдел тела Stereobalanus canadensis (Enteropneusta) (по Шпенгелю): 1 — спинной нервный тяж, 2 — гонады, 3 — кншка, 4 — жаберная дужка, 5 — брюшной нервный тяж, 6 — жаберное отверстие куолизированных клеток) и развития. Оба органа развиваются из части энтодермальных клеток, составляющих спинную стенку зародышевого кишечника. Нотохорд лежит в узком основании хоботка, прилегая к его брюшной стенке. Между эпителием брюшной стенки-хоботка и нотохордом развивается местное утолщение базальной мембраны, имеющее вид пластинки с двумя выростами, охватывающими с боков начало глотки. Это образование вместе с нотохордом придает хоботку известную плотность и укрепляет его основание. Глотка продолжается в пищевод, представляющий дыхательный отдел кишечника. Спинная и брюшная стенки пищевода сплошные, тогда как по бокам он пронизан двумя рядами жаберных щелей, сообщающих полость пищевода с внешней средой (рис. 556). Каждая жаберная щель имеет вид подковы, обращенной ветвями к спинной стороне тела, и представляет собой выпячивание стенки пищевода, подходящее к боковой стенке тела и там открывающееся наружу. Вода, заглатываемая через рот, выводится наружу через щели. В перегородках между щелями залегают кровеносные сосуды. В эти сосуды диффундирует кислород из воды, проходящей через жаберные щели. В перегородках между щелями развивается сложный скелет из вилообразных палочек, представляющих местные утолщения базальной мембраны. Этот скелет не дает стенкам щелей спадаться, что препятствовало бы прохождению воды. За жаберным отделом следует печеночный отдел кишечника с многочисленными боковыми кармашками железистого характера. Остальная часть средней кишки имеет вид простой трубки; недалеко от заднего 563 конца тела средняя кишка переходит в заднюю, которая и открывается анальным отверстием. Целом. Кишечник лежит во вторичной полости тела, разделенной на несколько участков. В воротничке и туловище 2 пары целомических мешков; мешки левой и правой сторон сходятся над и под кишечником, образуя спинно-брюшной мезентерий. Наружными стенками целомические мешки прилегают к стенке тела, внутренними — к кишечнику. Целом хоботка непарный; он довольно сильно редуцирован вследствие развития между его стенкой и мускулатурой толстого слоя соединительной ткани паренхиматозного характера. Кровеносная система развита довольно хорошо и состоит из брюшного и спинного продольных сосудов. Спинной сосуд через воротничок проходит в хоботок и, достигая задней стенки хоботкового целома, расширяется в кровеносную лакуну. Перегородка между лакуной и хоботковым целомом образует многочисленные складки (так называемый glomerulus), увеличивающие поверхность диффузии между обеими полостями. Это позволяет продуктам обмена веществ, скапливающимся в крови, диффундировать в целом хоботка и выводиться оттуда наружу через пору хоботка. Кровь течет в спинном сосуде вперед и на протяжении жаберного отдела туловища частично поступает в отходящие от сосуда парные приносящие жаберные сосуды. В стенках жаберных щелей сосуды распадаются на сеть лакун, где кровь окисляется, а затем по выносящим сосудам проходит в брюшной сосуд. Большая часть крови спинного сосуда, достигнув в хоботке центральной лакуны, направляется назад по двум окологлоточным сосудам, окаймляющим глотку в воротнике и впадающим в начало брюшного сосуда. По нему кровь идет к заднему концу тела, где через сеть околокишечных сосудов возвращается в спинной сосуд. Циркуляция крови по телу обусловливается присутствием особого пульсирующего органа — сердечного пузырька, или перикардия (рис. 555), исполняющего функции сердца, но не служащего для прохождения через него крови, подобно сердцу членистоногих и моллюсков. Это полый, замкнутый мускулистый мешок, лежащий у спинной стенки хоботка так, что центральная лакуна расположена как раз между ним и нотохордом. Мешок имеет мускульные волокна и способен с их помощью ритмически сокращаться и расширяться. При расширении сердечного мешка кровь, находящаяся в лакуне, между ним и нотохордом, выдавливается назад в окологлоточные сосуды. При сокращении мешка полость лакуны увеличивается и в нее поступает новая порция крови из спинного сосуда. Выделительная система устроена по типу целомодуктов. В более примитивных случаях она состоит из 2 пар коротких ресничных трубок, сообщающих целом хоботка и оба целома воротничка со средой. Судя по этому, целом хоботка первоначально был парным. У громадного большинства кишечнодышащих в хоботке сохраняется только один левый целомодукт — пора, открывающаяся наружу на спинной поверхности хоботка. Воротничковые целомодукты открываются не прямо наружу, а в первую пару жаберных щелей. Нервная система интересна тем, что кроме спинного нервного ствола имеется брюшной, но развитый значительно слабее. Брюшной ствол развит только на протяжении туловища; спинной — продолжается в воротничок и хоботок. У задней границы воротничка оба ствола соединены кольцевой комиссурой — воротничковым кольцом. Туловищные участки стволов, а также воротничковое кольцо залегают в самом эпителии, за- 564 нимая поверхностное положение. Напротив, спинной ствол воротничка лежит поверхностно лишь на молодых стадиях развития, позднее он впячивается сначала в виде желобка, который затем вполне отшнуровыва-ется от кожи и дает толстостенную нервную трубку. Просвет трубки позднее местами редуцируется. Эта часть нервной системы наиболее дифференцирована, и некоторые авторы даже называют ее «спинным мозгом». Кроме центральной нервной системы в коже рассеяно густое сплетение нервных клеток, отростки которых связаны с нервными стволами. Органы чувств отсутствуют. В наружном эпителии кишечнодышащих обнаружено большое количество светочувствительных клеток. Половая система устроена очень просто. Самцы внешне не отличимы от самок. Половые железы образуют свыше 30 пар овальных мешков, лежащих по бокам кишечника в средней части туловища и открывающихся наружу короткими выводными протоками на спинной стороне. Половые продукты выводятся в окружающую воду. 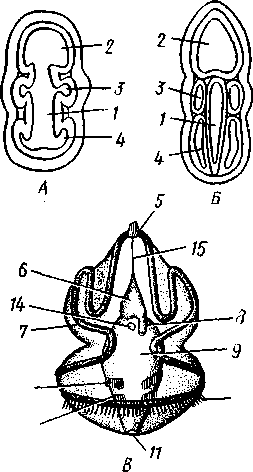 73 12 Ю Рис. 557. Развитие Enteropneus-ta. A—Б — энтсроцельный способ образования целомов у Ва-lanoglossus (по Бэтсону); В — торнария — личинка Enteropne-usta с брюшной стороны (из Мак-Брайда): / — кишечник, 2, 3, 4 — целомические мешки, возникающие в виде выступов кишечника и отшнуровы-вающиеся от него, 2 — целом хоботка, 3 — целом воротничка, 4 — целом туловища, 5 — теменная пластинка с теменным султаном, f> — передний целом (целом хоботка), 7 — ресничный шнур, 8 — поровый канал, 9 — средняя кишка, 10 — ресничный поясок, // — анус, 12 — задний целом (целом туловища), 13 — средний целом (целом воротничка), 14 — рот, /5 — мускульный тяж, соединяющий теменную пластинку с первым целомом Развитие. Полное, равномерное, радиальное дробление приводит к образованию бластулы, а затем типичной инвагинацион-ной гаструлы. Полюс гаструлы, противолежащий бластопору, отвечает будущему переднему концу животного, бластопорный полюс — заднему. На месте бластопора при дальнейшем развитии возникает порошица, а рот закладывается посредством вторичного впячивания эктодермы ^ла брюшной стороне личинки. Гаструла вытягивается в передне-заднем направлении, и начинается обособление мезодермы. Обособление происходит так называемым энтероцельным способом, т. е. при помощи образования кишечником переднего непарного и двух пар боковых выпячиваний, которые отшнуровы-ваются от энтодермы зародыша и превращаются в целомические мешки хоботка, воротничка и туловища (задняя, самая крупная пара боковых выпячиваний; рис. 557). У большинства кишечнодышащих имеется личинка торнария (рис. 557). За счет общего мерцательного покрова дифференцируются 2 венчика из крупных ресниц: околоротовой и околопорошицевый. Околоротовой венчик образует многочисленные извилины и загибы, вследствие чего торнария приобретает большое сходство с личинками иглокожих. На переднем полюсе торнарии образуется особое утолщение эктодермы — теменная пластинка с султаном ресниц и парой небольших глаз, позднее редуцирующихся. При дальнейшем развитии соответственно трем сегментам личинка слегка перетягивается на три отдела. Два передних отдела растут сравнительно слабо, тогда как задний удлиняется во много раз и дает ту- 565 ловище. Постепенно, по направлению спереди назад, прорываются жаберные щели. Молодое животное, плававшее до сих пор в планктоне при помощи своих ресничек, опускается на дно и переходит к роющему образу жизни. Экология и представители. Лишь немногие кишечнодышащие живут на поверхности грунта, большинство роет в нем ходы или делает постоянную норку, стенки которой укрепляются выделяемой животным слизью. Обычно норка имеет U-образную форму, оба конца ее открываются на поверхность грунта. У баланоглосса — Balanoglossus clavi-gerus переднее колено норки имеет несколько воронковидных входных отверстий — главный и запасные, а заднее выходное отверстие норки завалено колбасовидными экскрементами. Кишечнодышащие заглатывают грунт, а находящиеся в нем микроскопические организмы (фора-миниферы, водоросли и др.) и частицы детрита перевариваются в печеночном отделе кишечника. Обитают кишечнодышащие в теплых и холодных морях. В нашей фауне известен один вид из Белого и Баренцева морей, несколько других встречаются в дальневосточных морях. К данному классу относится несколько родов (Balanoglossus, Phychodera, Saccoglossus, Stereobala-nus и др.). Филогения кишечнодышащих. Долгое время класс Enteropneusta относили к червям, и только исследования А. О. Ковалевского (1867) показали, что они стоят гораздо ближе к хордовым животным. В общем можно сказать, что в строении взрослых Enteropneusta сказывается родство с типом Chordata (жаберные щели, спинная нервная трубка, нотохорд, сердечный мешок, гомологичный соответственному образованию у оболочников), но многие особенности развития живо напоминают иглокожих. Характер дробления, развитие порошицы на месте бластопора и вторичная закладка рта, энтероцельный способ закладки мезодермы общие для иглокожих и класса Enteropneusta. Кроме того, у иглокожих сходно закладываются 3 пары целомов, хотя в передней паре достигает развития лишь один левый; то же характерно и для кишечнодышащих, причем целомодукт хоботкового целома может быть приравнен каменистому каналу Echinodermata. Наконец, общий вид торнарии настолько похож на личинок иглокожих, что до 1870 г. ее считали личинкой какой-то морской звезды. КЛАСС II. КРЫЛОЖАБЕРНЫЕ (PTEROBRANCHIA) Современные крыложаберные представлены тремя родами — Atuba-ria, Cephalodiscus и Rhabdopleura, насчитывающими вместе 17 видов. Все они ведут сидячий образ жизни. В громадном большинстве это мелкие колониальные формы, образующие разветвляющиеся домики или трубочки; часто имеют вид кустиков. Таким образом, колония крыложа-берных напоминает мшанок, к которым их ранее и относили. Исключение представляет только род Atubaria; одиночные особи ее лишены трубочки и способны медленно ползать по субстрату. Строение отдельных особей во всех главных чертах напоминает класс Enteropneusta, а большинство отличий хорошо объясняется сидячим образом жизни. Тело состоит, как у кишечнодышащих, из трех сегментов, образующих три отдела: головной щит, воротничок и туловище (рис. 558). В головном щите имеется непарный целом, сообщающийся у всех форм с наружной средой двумя ресничными целомодуктами. В воротничке и туловище целомы парные; имеется пара воротничковых целомодуктов. Так же, как у кишечнодышащих, в основание переднего 566 отдела тела вдается нотохорд; со спинной стороны к нему примыкает сердечный мешочек. Между последним и нотохордом находится центральная лакуна спинного кровеносного сосуда. Мешковидные половые железы лежат в туловище. От кишечнодышащих отличаются укороченной формой тела, наличием на воротничке 2—12 перистых щупалец, покрытых ресничками, подгоняющих пищу ко рту, петлевидной изогнутостью кишечника и расположением порошицы на спинной стороне тела. Кроме того, у Rhabdopleura нет жаберных щелей, a Atubaria и Cephalo-discus имеют всего одну пару. Способность образовывать колонии связана с наличием бесполого размножения посредством почкования. Особой сложности достигают колонии Rhabdopleura (рис. 558), обнаруживающие в деталях строения, законах роста и в совершенно своеобразном характере почкования так много общего с колониями граптоли- 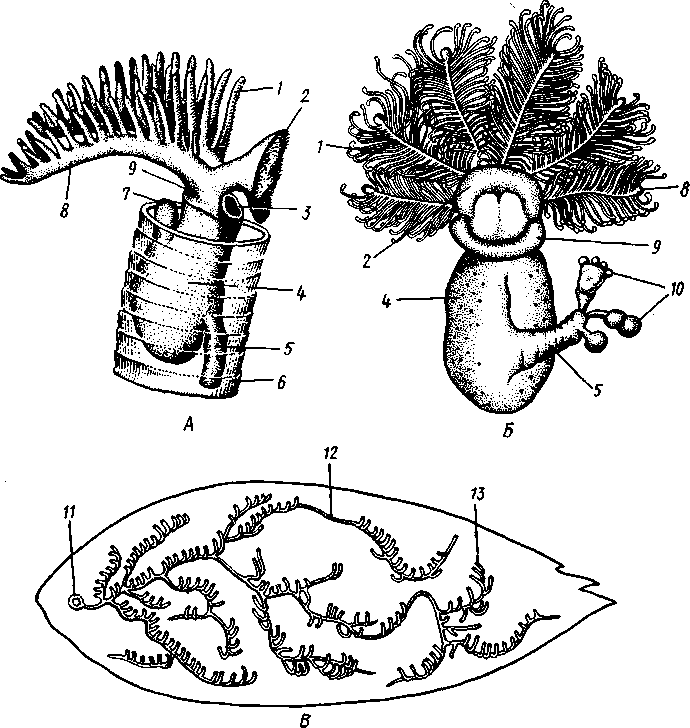 Рис. 558. Крыложаберные Pterobranchia. Л — зооид Rhabdopleura, вид справа (по Щепотьеву); Б — Cephalodiscus dodecalophus, вид с брюшной стороны (по Мак-Интош); В — колония Rhabdopleura на поверхности раковины моллюска (по Щепотьеву): / — выросты щупалец, 2 — головной щиток (первый сегмент), 3 — рот, 4 — туловище, 5 — стебелек, 6 — трубка, 7 — анус, 8 — щупальце — рука. 9 — воротничок, 10 — почки на стебельке, // — начальная часть колонии, 12 — столон, 13 — трубочки — ячейки отдельных зооидов 567 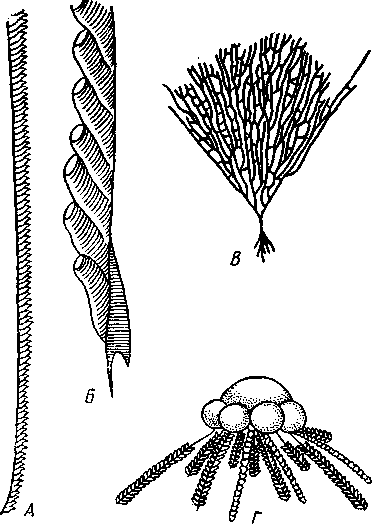 Рис. 559. Граптолиты Graptolithida. A — колония Monograptus; Б — она же, в увеличенном виде (по Рюдеманну); В — общий вид колонии Dictyonema (по Виману); Г — плавающая колония Diplograptus (по Рюдеманну) тов, что всякие сомнения в принадлежности последних к классу Pterobranchia отпадают. Граптолиты (Graptolithida) — богатая видами вымершая группа ', обитавшая в морях с кембрия до карбона, но особенно характерная для силура. Колонии граптолитов имели вид прямых изогнутых, иногда спиральных прутьев или небольших разветвляющихся кустиков, усаженных рядами ячеек, в которых помещались отдельные особи (рис. 559). Строение последних, вероятно, было очень близко к современным Pterobranchia. Колонии граптолитов могли прикрепляться ко дну или к плавающим водорослям, но в преобладающем большинстве плавали на поверхности моря вроде некоторых сифонофор. В связи с этим на одном конце колонии развивался плавательный пузырь, наполненный газом. ТИП ПОГОНОФОРЫ (POGONOPHORA) Погонофоры — тип целомических животных с небольшим числом видов (около 120), ставших известными науке лишь в текущем столетии. Первый представитель Siboglinum был описан в 1914 г. французским зоологом Коллери, второй— Lamellisabella — в 1933 г. П. В. Ушаковым, однако лишь в 1937 г. шведский исследователь Иоганессон показал, что эти животные относятся к новому классу, названному им Pogonophora. Основные особенности организации и эмбрионального развития погоно-фор были выяснены А. В. Ивановым (1955, 1960), установившим, что погонофоры являются самостоятельным типом. Для погонофор характерны следующие черты строения и развития: 1. Тело их состоит из четырех первичных сегментов, содержащих каждый свой отдел целома. Третий сегмент очень длинен, на его протяжении многие органы, в том числе участки целома, приобретают более или менее правильное метамерное расположение, проявляющееся в разных областях неодинаково. Четвертый сегмент отличается правильной вторичной сегментацией. 2. Кишечник полностью отсутствует, для собирания из воды пищевых частиц и для всасывания питательных веществ служат щупальца, развитые на первом сегменте. 1 Ряд исследователей рассматривают граптолитов как самостоятельный класс (Graptolitholitina) в составе типа полухордовых, другие включают их в класс крыло-жаберных. 568 3. Нервная система состоит из головного мозга и продольного нервного ствола, лежащих на брюшной стороне тела. 4. Имеется хорошо развитая кровеносная система, снабженная спинным сердцем, возле которого расположена околосердечная сумка (перикардий). 5. Органами выделения служат целомодукты первого сегмента. 6. Погонофоры раздельнополы. Половые железы лежат в туловищном сегменте, целомодукты которого служат для выведения половых продуктов. 7. Во время эмбрионального развития вторичная полость тела образуется энтероцельным путем. К типу Pogonophora относится один класс того же названия. |