А-Догель - Зоология беспозвоночных_2. Тип губки (spongia, или porifera)
 Скачать 14.77 Mb. Скачать 14.77 Mb.
|

|
Развитие прямое. Полное равномерное дробление приводит к образованию бластулы, а затем инвагинационной гаструлы. Зачатки половых желез закладываются на стадии 32 бластомеров (рис. 565). Еще в неоплодотворенном яйце имеется небольшое скопление особых зерен, называемое половым детерминантом. На стадии 32 бластомеров детерминант оказывается в одной из клеток. Дальнейшее деление этой клетки происходит так, что одна из дочерних клеток получает целиком весь де- 575 терминант и становится половым зачатком, тогда как другая лишена детерминанта и дает впоследствии всю энтодерму. Половой бластомер сначала делится надвое, а в гаструле половой зачаток имеет вид двух пар клеток, причем из одной пары возникают в дальнейшем яичники, из другой — семенники. Далее следует отметить энтероцельное образование мезодермы, отшнуровываюшейся от кишечника в виде двух боковых выпячиваний, полость которых представляет целом. Получаются два цело-мических мешка, из которых каждый перешнуровывается поперек на маленький головной и большой туловищный целомы. Последние позднее отделяют хвостовой целом. Бластопор гаструлы замыкается, и окончательно рот возникает независимо от него на противоположном конце зародыша. Филогения щетинкочелюстных. Тип Chaetognatha относится к вторич-норотым по способу закладки мезодермы и характеру возникновения рта. Однако тело щетинкочелюстных, судя по характеру его развития, слагается из двух сегментов, ибо перегородка, отделяющая полость хвоста от полости туловища, возникает значительно позже и носит вторичный характер. Этим тип Chaetognatha существенно отличается от вто-ричноротых, которые в основном трехсегментные животные. Таким образом, происхождение щетинкочелюстных еще совершенно не ясно и положение их в системе животных неопределенно. П 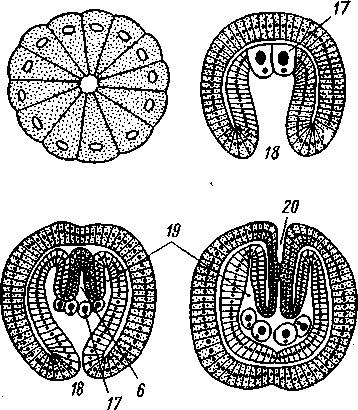 576 Рис. 565. Схема строения (А — вид с брюшной стороны, по Деляжу) и развития (Б — четыре последовательные стадии, по Р. Гертвигу) Sagitta: I — мозг, 2 — окологлоточный коннектив, 3 — брюшной ганглий, 4 — нервы, 5 — целом, 6 — кишка, 7 — яичник, 8 — яйцевод, 9 — анус, 10 — семенник, // — семяпровод, 12 — хвостовой плавник, 13 — головная железа, 14—хи-тиноидные крючки — щетинки, 15 — мужское половое отверстие, 16 — женское половое отверстие, 17 — первичные половые клетки, 18 — бластопор, 19 — боковые выпячивания первичной кишки — зэчатки целома (энтероцельный способ образования целома), 20 — вторичный рот ОСНОВНЫЕ ЭТАПЫ ФИЛОГЕНЕТИЧЕСКОГО РАЗВИТИЯ ЖИВОТНОГО МИРА В настоящей главе мы дадим краткий обзор усложнения организации беспозвоночных, возникающего в процессе эволюции, не останавливаясь на родственных отношениях отдельных групп. Филогения отдельных групп кратко разобрана в соответствующих местах учебника. Рассматривая огромное разнообразие форм беспозвоночных, можно по степени сложности строения расположить эти формы на нескольких уровнях. В главных чертах это ярусное расположение отвечает эволюционному пути, пройденному животными. На филогенетическом древе ярусы могут быть отделены друг от друга горизонтальными линиями. Группы животных, расположенные между одними и теми же горизонталями, имеют приблизительно одинаковую сложность организации (см. табл. на с. 578). Первый этаж родословного древа образован полцарством Protozoa, включающим пять типов. Мы уже видели, что в разных типах Protozoa (Sarcomastigophora и др.) обнаруживается тенденция к переходу в многоклеточное состояние (многоядерность, образование колоний и т. п.). Эта тенденция большей частью остается незавершенной, но у каких-то Mastigophora привела к прочному переходу к многоклеточное™: возникли первые Metazoa. Низшие Metazoa образуют следующие этажи родословного древа. К ним принадлежат четыре типа Placozoa: Spongia, Coelenterata и Cte-nophora. Из них губки находятся на гораздо более низком уровне развития, о чем прежде всего свидетельствует отсутствие у них настоящих тканей, а также нервной системы. Кроме того, губки существенно отличаются от других Metazoa «извращением» положения зародышевых листков. В связи с этим следует считать, что Spongia отделились от общего ствола Metazoa чрезвычайно рано, вероятно еще на уровне фаго-цителлы — гипотетического организма, который, по мнению И. И. Мечникова, был общим предком всех многоклеточных животных. Таким образом, губки составляют среди Metazoa особый ярус — надраздел Ра-razoa, противопоставляемый всем остальным многоклеточным — надраз-делу Eumetazoa. Coelenterata и Ctenophora образуют следующий ярус — раздел Radiata. Общность особенностей строения позволяет эти два типа именовать Radiata или Diploblastica. Характеризуются они тем, что тело их состоит из производных двух зародышевых листков — эктодермы и энтодермы, которые даже у взрослых животных сохраняют свою обособленность и характерное расположение. По плану строения они сходные гаструлой и обладают радиальной симметрией. Можно предположить, что первичные Coelenterata произошли от фагоцителлы, причем Ctenophora сохранили древний способ движения — плавание посредством ресничного аппарата, a Coelenterata перешли к сидячему образу жизни. В обоих случаях сохранилась первобытная радиальная симметрия тела. 20—763 577 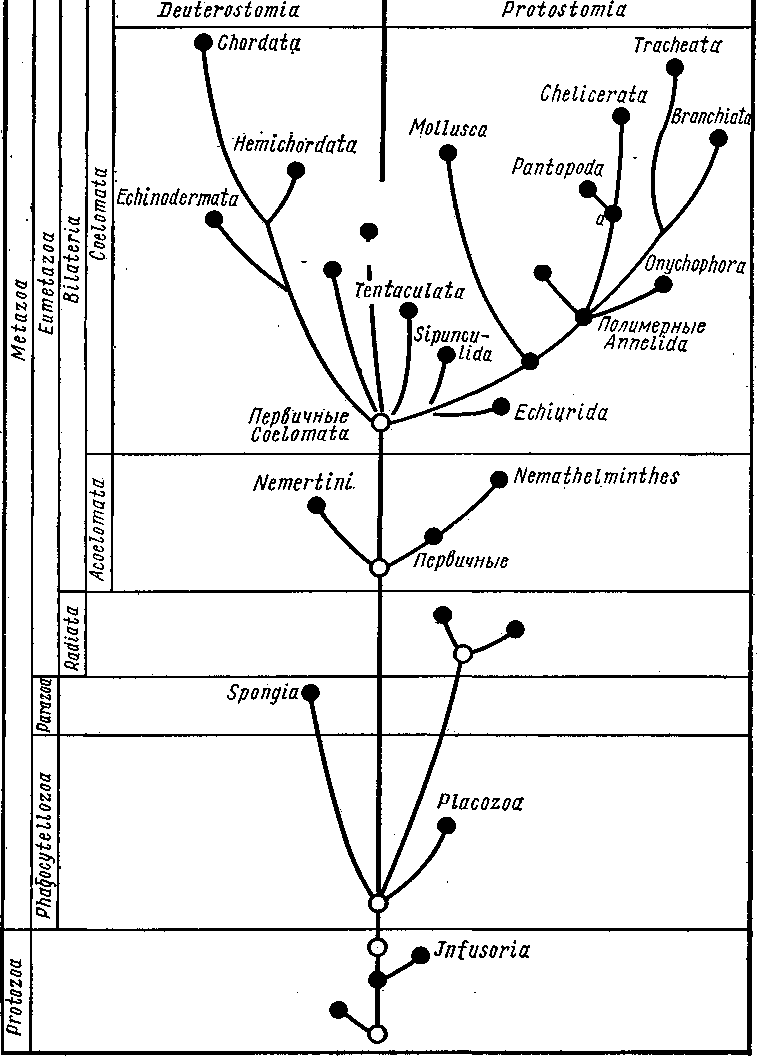 Chaetognatha Poaonophora. Trilobitomorph Tardlgrada 0/шгомернь/е Annelida Plathelmintties ацелооУразные Bilaterlu Ctenophora Coelenteraia Фагоцителла Колониальные Mastigoptiora. Mastigophora Sarcodim Первичные эукариотные простейшие Все вышестоящие Eumetazoa в отличие от Radiata характеризуются билатеральной симметрией тела и объединяются в раздел Bilateria. Другое их название Triploblastica, так как тело их формируется в онтогенезе из трех зародышевых листков — экто-, эндо- и мезодермы. Ни губки, ни кишечнополостные не дали начала новым типам животных. По-видимому, Bilateria произошли независимо от тех и других, непосредственно от фагоцителлообразных предков, перешедших к ползающему образу жизни, вызвавшему появление билатеральной симметрии. Наиболее примитивные Bilateria в мире животных представлены свободноживущими плоскими червями — Plathelminthes, именно классом ресничных червей Turbellaria. Среди последних наиболее примитивные представители — отряд бескишечных Acoela. По принципу организации, а для них характерна центральная масса клеток пищеварительной паренхимы (фагоцитобласт), бескишечные турбеллярии напоминают еще фагоцителлообразных прародителей Metazoa. Низшие Bilateria, в первую очередь плоские черви (тип Plathelminthes), составляют пятый ярус родословного древа животного мира. Билатеральность, централизация нервной системы, выражающаяся в появлении головного мозга, дифференциация осморегуляторной и выделительной системы (протонефридиев), ясная дифференцировка мезодермы во время развития ставят их на более высокую ступень, чем Radiata. К этому же ярусу кроме плоских червей принадлежат типы Namathel-minthes и Nemertini. Одним из очень характерных общих признаков всех перечисленных типов следует считать отсутствие у них вторичной полости тела, что позволяет под названием Acoelomata противопоставлять их всем высшим Bilateria, для которых, напротив, характерно развитие це-лома (Coelomata). Как синоним Acoelomata некоторыми зоологами употребляется название Scolecidae — низшие черви. Все крупные группы сколецид, вероятно, произошли от ресничных червей или от очень близких к ним, не сохранившихся турбелляриеобразных предков. Верхний ярус филогенетического древа составляют вторичнополост-ные животные — Coelomata. Среди них имеются как вероятно первично несегментированные формы с общим неметамерным целомом (Echiurida, Sipunculida), так и животные, низшие представители которых имеют ясную метамерию наружных органов, но сохраняющие несегментированный целом (тип Mollusca). Последние, вероятно, произошли от примитивных олигомерных кольчатых червей, обладающих уже наружной метамерией, но лишенных еще сегментации вторичной полости тела. Среди современных кольчецов такие близкие к общему предку Coelomata формы есть. Таков, например, Dinophilus (с. 252), не обладающий еще пара-подиями и щетинками, а также кровеносной системой и состоящий из 5 ларвальных сегментов. По плану строения Dinophilus близок к мета-трохофоре; целом у него не подвергается сегментарному расчленению, что характерно также для многих метатрохофор. Большинство Coelomata, однако, составляют сегментированные животные с метамерно расчлененным целомом. Среди последних различают малосегментные формы (типы Tentaculata, Hemichordata) и многосегментные, или полимерные (типы Annelida, Arthropoda, Chordata). Происхождение Coelomata неясно и спорно. Наиболее вероятной кажется нам гипотеза, по которой их предками считаются турбелляриеобразные животные, давшие начало также современным турбелляриям и немертинам. Основная черта, определяющая более высокую организацию Coelomata,— возникновение вторичной полости тела. Их эволюция сопровождалась развитием кровеносной системы, превращением протонефридиев в метанефридии, формированием целомодуктов. Таким образом, органи- 20* 579 зация животных при переходе к Coelomata подвергалась серьезной перестройке, носившей прогрессивный ароморфный характер. Следует, однако, заметить, что некоторые из вновь возникших систем органов остаются у высших червей еще слаборазвитыми. Так, в кровеносной системе обычно отсутствует центральный пропульсаторный орган — сердце (лишь у некоторых плеченогих имеется образование, напоминающее сердце). Почти то же можно сказать о дыхательной системе, органы которой очень часто (тип Tentaculata, часть представителей многощетинковых кольчецов Annelida) наряду с дыхательной несут иную функцию (захват пищи, чувствительная), т. е. не строго специализированы. Типы Arthropoda и Mollusca берут начало от кольчатых червей: первые от полимерных, вторые от олигомерных. Оба типа представлены вторичнополостными животными, у которых заложившиеся на предыдущем этапе филогенетического развития органы достигают значительной степени совершенства. Кровеносная система приобретает сердце, органы дыхания (жабры р?,ков, ктенидии моллюсков, трахеи насекомых) строго специализируются и служат исключительно для газового обмена. Большой шаг вперед делает нервная система: центральная и периферическая. В центральной нервной системе у многих представителей обоих типов наблюдается, с одной стороны, сильная концентрация ганглиев, а с другой — увеличение относительных размеров мозга. Поражает сильное развитие органов чувств, особенно глаз, которые у головоногих, высших раков и насекомых не уступают в сложности строения органу зрения человека. Располагая моллюсков и членистоногих в одном горизонтальном ярусе, мы несколько затруднены в оценке того, который из этих типов более высоко организован. С одной стороны, моллюски имеют более совершенную кровеносную систему: это единственные из беспозвоночных, обнаруживающие дифференциацию сердца на желудочек и предсердие. С другой стороны, членистоногие снабжены несравненно более совершенными органами движения: богато расчлененные, способные к очень сложным движениям конечности, а кроме того, крылья. Эти свойства, равно как и развитие высшей нервной деятельности (у общественных насекомых в особенности) и воздушных органов дыхания, привели тип Arthropoda в лице высших его представителей, т. е. паукообразных и насекомых, к широкой адаптации к наземному образу жизни. Кроме позвоночных, мы ни у одной группы животных не встречаем таких многочисленных и разнообразных приспособлений к разным условиям сухопутного существования. Среди моллюсков выход на сушу наблюдается несравненно реже и не влечет к столь пышному расцвету форм, какой наблюдается у перешедших к наземной жизни представителей типа Arthropoda, а именно у насекомых. Обсуждая нашу схему родословного древа, мы говорили до сих пор лишь о горизонтальных разделениях — ярусах. Но кроме того, можно провести вертикальную линию, рассекающую Coelomata на две большие группы: первичноротые—Protostomia и вторичноротые — Deutero-stomia. Очевидно, эти филогенетические ветви имели общее начало где-то среди первобытных Coelomata, а затем развивались самостоятельно и параллельно друг другу. Возможно, что некоторые примитивные группы Coelomata, в первую очередь Sipunculida и Tentaculata, в какой-то мере являются промежуточными между первичноротыми и вторичноротыми, и мы лишь условно относим их к Protostomia. 580 Главные различия между Protostomia и Deuterostomia эмбриологического порядка уже приводились (с. 289). Ход эволюции Protostomia нами только что рассмотрен. Исследования последних лет показывают, что кроме двух основных эволюционных путей развития Coelomata: первичноротых и вторичноро-тых, есть и другие независимые пути эволюции, которые, возможно, берут начало от неизвестных нам общих предков Coelomata. Такими независимыми линиями развития следует считать щупальцевых (Tenta-culata), щетинкочелюстных (Chaetognatha) и погонофор (Pogonophora) (см. схему с. 578). Последние обладают четырьмя сегментами (а не тремя, как полагали еще недавно), что не позволяет непосредственно включать погонофор во вторичноротых, с которыми они обнаруживают многие черты сходства (в том числе энтероцельный способ образования целома). На организацию погонофор сильный отпечаток наложило обитание в длинных защитных трубках, погруженных нижним концом в ил. Эволюция Deuterostomia гораздо менее ясна, чем Protostomia. Тип Echinodermata в смысле высоты организации стоит приблизительно уже на уровне высших червей. В самом деле, наличие целома и кровеносной системы уравнивает иглокожих с высшими червями. Дыхательная система иглокожих не строго специализирована, как и у высших червей, ибо кожные жабры их большей частью несут также иные функции: например, через жабры идут процессы выделения (выход амебоцитов) и т. д. Кровеносная система еще не централизована, а нервная находится даже на низшей ступени централизации, чем у кольчатых червей. Второй, относящийся ко вторичноротым, тип Hemichordata также стоит на низшей ступени развития этой ветви животного царства. Организация обоих типов сводится к трехсегментному строению, столь характерному для диплеврулы, в которой естественно видеть рекапитуляцию общего предка всех вторичноротых. У Hemichordata наблюдается явный переход к полимерному состоянию путем вторичной метамеризации туловищного сегмента. У вторичноротых, следовательно, также можно различать ларваль-ные и постларвальные сегменты. Однако полимерность у них достигается совершенно иным способом, чем у первичноротых. У Deuterostomia постларвальные сегменты представляют многочисленные метамерные участки сильно вытянутого заднего (третьего ларвального) сегмента, тогда как у Protostomia они всегда образуются на заднем конце тела как добавление к последнему ларвальному сегменту. Таким образом, постларвальные сегменты первичноротых и вторичноротых не гомологичны друг другу. Наконец, третий тип вторичноротых, а именно Chordata, находится на уровне высших Protostomia и даже поднимается выше. Нервная система хордовых достигает степени дифференциации, неизвестной среди беспозвоночных. То же следует сказать об органах слуха, тогда как глаза хордовых не превышают по сложности строения глаза высших беспозвоночных, а именно головоногих моллюсков. Кровеносная система достигает высшего во всем животном мире развития, хотя у низших представителей хордовых она устроена просто. Выделительная система обнаруживает характер целомодуктов, но чрезвычайно усложнена. Дыхательная система строго специализирована. Усложнение строения хордовых обусловлено тем, что эта группа перешла от водного к наземному образу жизни (подобно классам Arachni-da и Insecta среди Arthropoda). Указанные признаки свидетельствуют, что тип Chordata должен быть отнесен к высшему из намеченных нами этапов усложнения. 581 УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ И ТЕРМИНОВ 321, 322, 338, 339, 432, 437, 296, 297, 312, 313, 320, 321, 404, 412, 403, 404, 448, 451, 404, 416, Абиссаль, 5, 22 Аборальная система 526*, 528 Аборальная сторона 524, 529 Аборальный орган 145 Аборальные пластинки 532 Авикулярия 503, 507, 508* Австралийская область 6 Автогамия 70, 79 Агамета 30 Агамонт 30 Адамбулакральные пластинки 524, 525* Адаптивная радиация 8 Аденофореи 228*. 229 Адолескария 171 Аккомодация 493 Акрон 291, 293, 295, 323, 326, 331, 332, 333, 339, 341, 354, 399, 401, 402, 406, 412, 425, 437 Аксоподии 32, 35 Аксостиль 51 Актинии 137, 141 Актиномиксидии 67 Актинотроха 518, 519* Альционарии 140 Амбулакральная борозда 524 Амбулакральная система 523, 528* Амбулакральные ножки 524, 529 Амбулакральные пластинки 524, 529, 535 Амебоидный зародыш 70 Амебоидные клетки 287 Амебоцит 103, 279 Амебиаз 26 Амебы 23, 24 Амитоз 26 Аммониты 497, 498, 499 Амнион 370 Амфибии 51 Амфибластула 109 Амфиды 214, 216* Амфилина 197*, 198 Анабиоз 240 Анальная лопасть 250, 267 Анальные мешки 283, 284 Анаморфоз 338, 371, 388, 434 Анаэробный обмен 173, 183, 202 Анизогамия 45, 46, 52, 55 Анимальное питание 40 Анимальные клетки 261 Аннелиды 285, 436 Антарктическая область 6 Антенна 253, 259, 260, 293, 294, 295, 296, 297, 303, 306, 308, 311, 312, 313, 314, 315, 316, 317, 318, 323, 324, 331, 333, 334, 336, 340, 341, 374, 400, 427, 431, 438, 439 Антеннула 291, 293, 294, 295, 303, 304, 306, 308, 309, 311, 314, 315, 316, 317, 318, 319, 322, 323, 324, 331, 399, 401, 430, 435, 437 Аорта 292, 301, 337, 365, 366, 415, 416, 444, 445, 447, 442, 460 Аргонавт 499 Аристотелев фонарь 537, 540 Арканчик 128*. 129 Ароморфоз 8 Артерия 292, 300, 301, 337, 403, 447, 454 Асимметрия 453, 465 Аскарида 212*, 213*, 215* Аскон 102 Аспидогастры 175 Атокная часть тела 261* Атолл 139 Аттрактанты половые 359 Аурикулярия 555*, 556 Археоциты 103, 108 Аутотрофное питание 41 Бабочки 394 Бадяга 108* Базальная мембрана 119, 150 Бактриты 497 Баланоглосс 566 Барьерный риф 139 Батиаль 5 Бедро 345, 346 Беззамковые мшанки 512, 517 Беззубка 473* Безногие 543, 547 Белемниты 497, 498, 499 Белянка капустная 395 Бентос 5, 22, 272 Береговой риф 139 Бескишечные 158 Беспанцирные 449 Беспоясковые 250 Бессяжковые 387 Бивиум 543 Билатеральные 148 Бинарная номенклатура 12, 17 Биогенетический закон 11 |