биополимеры. Титульник Содержание
 Скачать 64.11 Kb. Скачать 64.11 Kb.
|

|
Титульник Содержание: Введение…………………………………………………………………………...3 1. Характеристика полисахаридов, получаемых путем биосинтеза (экзо – и эндополисахаридов)……………………………………………………………..5 2. Микроорганизмы – продуценты полисахаридов……………………………..6 3. Общие сведения о технологиях культивирования продуцентов полисахаридов…………………………………………………………………….8 4. Технология получения декстрана……………………………………………16 5. Технология получения ксантана……………………………………………..20 6. Технология получения альгината…………………………………………..21 Заключение……………………………………………………………………….22 Список литературы……………………………………………………………...23 Введение Термин «биополимеры» относится ко многим высокомолекулярным соединениям (полисахаридам, липидам, полиоксиалканоатам), которые являются для клеток резервными веществами и синтезируются в специ-фических условиях несбалансированного роста. Такими условиями, как правило, являются избыток углеродного и энергетического субстратов в среде и дефицит отдельных минеральных элементов (азота, фосфора, серы, магния и т.д.), лимитирующих синтез азотсодержащих компонентов и скорость роста клеток. Многие микробные биополимеры являются эндо-генным источником углерода и энергии, поэтому способствуют сохранению выживаемости клеток в неблагоприятных условиях среды. Выделяют два типа биополимеров — регулярные (некоторые полисахариды) и нерегулярные (белки, нуклеиновые кислоты, некоторые полисахариды) Полисахариды (гликаны) – полимеры, построенные не менее чем из 11 моносахаридных единиц. Полисахариды являются обязательным ком-понентом всех организмов, присутствуют как изолированно, так и в ком-плексах с белками, липидами, нуклеиновыми кислотами. Полисахариды преобладают в растительных биомассах и составляют, следовательно, большую часть органического материала на планете. Полисахариды раз-нообразны по строению, локализации в клетках и, естественно, по своим физико-химическим свойствам. Особенно разнообразны полисахариды, синтезируемые микроорганизмами. Микробные полисахариды делятся на внутриклеточные, локализованные в цитоплазме, и внеклеточные – поли-сахариды слизей, капсул, чехлов. Многие полисахариды биологически активны и повышают устойчивость макроорганизмов к вирусной и бакте-риальной инфекциям, обладают противоопухолевым действием, а также антигенной специфичностью. Поэтому они находят все более широкое применение в медицине и фармацевтической промышленности в качестве диагностикумов, заменителей плазмы крови и пр. Чрезвычайно широки перспективы применения полисахаридов в связи с их гелеообразующими и реологическими свойствами в качестве загустителей сиропов и космети-ческих средств, для упаковки продуктов и протравливания семян. Водные растворы отдельных полисахаридов чрезвычайно стабильны в широких интервалах рН и температуры, поэтому находят применения при добыче нефти и газа; флоккулирующие свойства гликанов используют в процессах очистки, концентрирования и разделения металлов. Возможности полисахаридов раскрыты далеко не полностью, поэтому их изучение ведет к расширению сферы применения Сферы использования полисахаридов: - медицина; - пищевая промышленность; - микробиология; - химия; - сельское хозяйство. 1 Характеристика полисахаридов, получаемых путем биосинтеза (экзо – и эндополисахаридов) Полисахариды микроорганизмов в соответствии с локализацией делятся на внутриклеточные и внеклеточные. К внутриклеточным относят обычно полисахариды цитоплазмы, мембран и клеточных стенок, а к внеклеточным – полисахариды капсул, чехлов (пристеночные структуры) и в свободной слизи, не прилегающей к клеточной стенке. Иногда к внеклеточным относят также полисахариды, локализованные снаружи от цитоплазматической мембраны. В этом случае в группу внеклеточных попадают и полисахариды клеточных стенок. У ряда микроорганизмов действительно трудно различить границу между капсулой и клеточной стенкой. Нередко по локализации выделяют три группы полисахаридов: внутриклеточные (цитоплазмы, мембран, периплазмы), полисахариды клеточных стенок и внеклеточные (капсул, чехлов и свободной слизи). Термин «экзогликаны» применяют в основном к полисахаридам свободной слизи. Иногда экзогликанами называют также капсульные полисахариды. Микробные полисахариды объединяют в группы и по функциям: резервные, участвующие в активном транспорте, опорные, участвующие во взаимодействии между клетками, защитные и др. Некоторые исследователи классифицируют полисахариды, учитывая их топологию и функции. В соответствии с этим клеточные полисахариды подразделяются на две группы. Одна включает резервно-энергетические и модификаторы (внутриклеточные), вторая – структурные и структурно-метаболические (в клеточной стенке). К внеклеточным относятся выделяющиеся при гиперпродукции структурно-метаболические гликаны и собственно гликаны. 2 Микроорганизмы – продуценты полисахаридов Полисахариды цитоплазмы обнаруживаются в двух формах: они могут быть диспергированы в ней или объединены в гранулы. Обычно в цитоплазме бактерий содержится 20-30% полисахаридов, а в условиях, способствующих их накоплению, до 50-60% от массы сухих клеток. Чаще всего в цитоплазме микроорганизмов обнаруживают гомогликаны, из которыз особенно распространены глюканы типа гликогена. Их выделяли из цитоплазмы многих эукариотных и прокариотных микроорганизмов: представителей разных родов бактерий Agrobakterium, Arthrobakter, Bakillus, Clostridium, Escherichia, Mykobakterium, Nostoc, Rhodopseudomonas, Rhodospirillum, Streptococcus, а также дрожжей, мицелиальных грибов, ресничных и жгутиковых простейших, некоторых водорослей. Кроме гликогеноподобных полисахаридов в цитоплазме ряда микроорганизмов найдены крахмал, маннаны, леваны, арабаныи ксиланы. Гликоген и другие гомогликаны цитоплазмы могут образовывать комплексы с ДНК, РНК, белками, липидами, фосфатом. Гетерополисахариды обнаруживаются в цитоплазме реже. Однако у представителей Streptomyces и Mycobakterium они оказываются преобладающими. У грамположительных эубактерий полисахариды составляют от 30 до 60% сухой массы клеточной стенки. Значительная их часть входит в состав муреинового комплекса, количество которого у грамположительных эубактерий достигает 50-90% веществ клеточной стенки. Линейные полисахариды в цепи муреина построены из повторяющихся β-1,4- связанных единиц N – ацетилглюкозамина и N- ацетилмурамовой кислоты. Мурамовая кислота – производное глюкозамина, содержащее D- молочную кислоту. Полисахариды – главные компоненты клеточной стенки дрожжей и мицелиальных грибов. Они могут составлять до 90% массы клеточной стенки (Saccaromyces cerevisiae). У дрожжей часто обнаруживаются гомогликаны: глюканы, маннаны, хитин. Внеклеточные полисахариды, капсульные или свободные, или те и другие, образуют многие микроорганизмы. Пожалуй нет такой группы микроорганизмов, представители которой не обладали бы этой способностью. Внеклеточные полисахариды большинства видов бактерий – кислые гетерогликаны разнообразного состава, построенные из 2-5, иногда 6-7 мономеров, линейные и разветвленные, имеющие регулярную структуру из повторяющихся олигосахаридных звеньев. Так, например, Xanthomonas campestris синтезирует полианионит ксантан, включающий глюкозу, маннозу, глюконовую кислоту и О – ацетильную группу и пируват. Наиболее активные продуценты декстранов – представители молочнокислых бактерий Leuconostoc mesenteroides и L.dextranicum. Декстраны синтезируются также некоторыми видами Streptococcus (Str.sanguis, Str. mutans), Brevibacterium, Lactobacillus. Практически каждый продуцент синтезирует свой, несколько отличный от других видов декстран. Некоторые штаммы образуют одновременно два различных по структуре декстрана. Внеклеточную целлюлозу – полисахарид, из бактерий синтезируют некоторые представители Pseudomonas, Zooglea, Azotobakter, подавляющее число видов Rhizobium и Agrobakterium, Acetobakter xylinum. Известно, что не только разные виды одного рода микроорганизмов, но и часто разные штаммы одного вида синтезируют неодинаковые экзополисахариды. Так, E. Coli имеет около70 серотипов, различных по составу и структуре капсульных полисахаридов и, соответственно, по иммунохимическим свойствам. Экзогликаны эффективных и неэффективных штаммов клубеньковых бактерий различаются по мономерному составу, а разных штаммов дрожжей рода Lipomykes – по соотношению моносахаров. Неодинаковыми по моносахаридному составу могут быть внеклеточные гликаны М-, S- и R- форм бактерий. Отмечены случаи, когда в культуральной жидкости одного микроорганизма накапливается несколько различных гликанов. Таблица 1. Промышленные микробные полисахариды
3 Общие сведения о технологиях культивирования продуцентов полисахаридов Большинство микроорганизмов синтезируют полисахариды из разнообразных источников углерода, обеспечивающих их рост, – углеводов, спиртов, карбоновых кислот, С1-соединений. Природа и концентрация углеродного источника в среде существенно влияет на образование полисахаридов, которое сводится к созданию гликозидной связи между моносахаридными единицами; при этом гликозильный донор передает гликозил на акцептор-затравку, высвобождаясь при этом. Акцепторами служат олигосахара и недостроенные полисахариды. Часто первичным акцептором служат олигосахара, в ряде случаев – недостроенный полисахарид – «затравка». Полимеризация идет до образования готового полисахарида с участием специфических гликозилтрасфераз, которые отщепляют фрагменты линейной цепи недостроенного гликана и переносят их на ту же или аналогичную цепь в определенном положении. Синтез полисахаридов определяется условиями культивирования продуцента и составом питательной среды, которые определяют возможность и интенсивность их образования, а также состав, структуру и, следовательно, свойства. Существенное значение имеют не только качественный состав используемого углеродного сырья, но также и концентрация, так как эффективный синтез полисахаридов осуществляется на средах с высоким содержанием углеродного субстрата. Количество и форма источника азота, не влияя на состав полисахаридов, оказывает влияние на скорость роста микроорганизмов и количественных выход полисахаридов. Существенна также роль фосфатов и ионов марганца, магния, кальция, являющихся кофакторами синтеза полисахаридов. Разнообразно и специфично влияние рН и температуры среды на накопления гликанов. Существенен хороший уровень аэрации культуры. Производство полисахарив специфично для каждого и определяется природой, локализацией, свойствами, а также областью применения гликанов и, безусловно, физиологическими особенностями продуцента. Получение экзополисахаридов эффективнее внутриклеточных, так как их концентрация выше, меньше проблем на стадии выделения и очистки, однако в ходе ферментации возникают трудности с транспортом кислорода из газовой фазы в жидкую (при повышении экскреции гликанов в среду ее вязкость возрастает). Следствием этого становятся снижение роста клеток и торможение продукции полисахаридов. Поэтому среду приходится разбавлять в десятки раз и после удаления клеток продуцента – концентрировать. Полисахариды, образуемые микроорганизмами, входят в состав ряда компонентов клеток, в частности в клеточную стенку, служат запасными веществами, локализуются внутри и вне клетки. Внеклеточные полисахариды могут быть капсульными или свободными. Часто полисахариды определяют антигенную специфичность штамма, вполняют защитную функцию, играя роль барьера проницаемости для молекул, ионов или препятствуют потере клеткой влаги. Из культуральной жидкости полисахариды выделяют осаждением 3—5 объемами этанола или изопропанола, осадок, образующийся за 24 ч при комнатной температуре отделяют центрифугированием (3 тыс. g, 15 мин) и сушат на воздухе. Для определения моносахаридного состава проводят гидролиз 1—2%-ного препарата полисахарида в 1 н. НС1 при 100° в течение 3 ч в герметическом сосуде и смесь затем доводят сухим бикарбонатом натрия до нейтральной реакции по индикатору. Раствор гидролизованного полисахарида хранят в холодильнике. Количество редуцирующих сахаров в гидролизате полисахарида определяют по восстановлению 2,3,5-трифенилтетразолия хлористого (ТТХ), используя в качестве стандарта раствор D-глюкозы. Качественный моносахаридный состав полисахарида определяют методом хроматографии на бумаге с соответствующими свидетелями. Для определения образования внутриклеточных полисахаридов в качестве запасных веществ проводят кислотный гидролиз клеточной массы и в гидролизате определяют количество редуцирующих сахаров. Нередко нативные полисахариды не обладают желаемыми качествами. Они могут быть, например, недостаточно активны или, будучи весьма высокоэффективными, плохо растворимы или оксичны, что препятствует их применению, и тд. Чтобы улучшить действие полисахаридных препаратов, устранить или снизить нежелательные явления, т.е. получить препарат с нужными свойствами, выделенные полисахариды иногда подвергают химической модификации. Так, декстран сульфатируют, чтобы придать ему антикоагулирующую активность. Обработка нативного гликана А.faekalis var. Myxogenes солюбилизирующими агентами позволяет получить производные, образующие гели без предварительного нагревания. Растворимость глюкана А.pullulans и маннана R.rubra удается повысить карбоксиметилированием. Снижение токсичности противоопухолевого препарата белково-липо-полисахаридного комплекса из культуральной жидкости Serratia piscatorum достигается обработкой его щелочью. Среда обитания микроорганизмов часто определяет количест венный выход, мономерный состав, структуру и молекулярную массу полисахаридов, а нередко — и самую возможность их био синтеза. Изменение условий культивирования сказывается в первую очередь на образовании внеклеточных и запасных цитоплазматических гликанов. Количество и состав структурных по лисахаридов отличаются сравнительным постоянством. Вследст вие большого разнообразия микробных гликанов и физиолого-биохимических особенностей их продуцентов значимость одного и того же фактора для образования различных полисахаридов не всегда одинакова. Состав питательной среды Источник углерода. Большинство микроорганизмов синтези рует полисахариды из всех источников углерода, обеспечиваю щих их рост: углеводов, спиртов, карбоновых кислот, амино кислот, углеводородов, Сi-соединений. В настоящее время особое внимание привлекает возможность образования микроорга низмами полисахаридов на средах с углеводородами и Ci-соединениями. На средах с н-алканами экзополисахариды синтези руются многими представителями артробактерий и сапротрофных микобактерий, некоторыми коринебактериями, дрожжами рода Candida. Ряд метилотрофных бактерий образует значитель ные количества внеклеточных полисахаридов при росте на сре дах с метанолом. Methylococcuscapsulatusобразует полисахаридные капсулы и при использовании метана. Помимо микроорганизмов, синтезирующих полисахариды на средах с любым источником углерода, допускающим рост, встре чаются организмы, образующие гликаны лишь при использова нии некоторых из них. Например, Leuconostocmesenteroidesможет расти, потребляя различные углеводы, но синтезирует декстран только на средах с сахарозой. Природа соединения углерода и его концентрация могут влиять на количественный выход полисахаридов. Так, наиболее высокий выход экзогликана в культурах некоторых микобактерий наблюдается в среде с гексадеканом, хотя биосинтез поли мера возможен на средах с другими углеводородами, сахарами и многоатомными спиртами. В ряде случаев для максимального образования полисахарида требуется более высокая концентра ция источника углерода в среде, чем для наивысшего накопления биомассы. В условиях периодического культивирования син тезу полисахаридов обычно благоприятствует создание избытка углерода в среде при некотором дефиците азота и фосфора. Однако стимуляция биосинтеза экзогликанов повышением содер жания углеродного субстрата в среде возможна лишь до опре деленного предела, сверх которого положительный эффект не проявляется, а в некоторых случаях наблюдается угнетение про цесса. При культивировании в хемостате некоторые микроорга низмы образуют внеклеточные гликаны даже при лимитации углеродным субстратом. Моносахаридный состав гликанов микроорганизмов не меня ется в зависимости от источника углерода, варьируют только состав и соотношение неуглеводных компонентов. Если же мик роорганизм синтезирует более одного экзогликана, изменение углеродного субстрата часто ведет к преимущественному синтезу одного из полимеров. Например, соотношение аубазидана и пуллулана в культуре Л, pullulansсильно различается на средах с разными сахарами. Изменение концентрации источника угле рода может регулировать скорость синтеза полисахаридов и их молекулярную массу. Источник азота. Образование полисахаридов возможно, как правило, при использовании источников азота, способных под держивать активный рост продуцента. Но природа источника азота может, не изменяя рост микроорганизма, влиять на коли чественный выход глкканов. Существуют микроорганизмы, для роста которых предпочтительнее один источник азота, а для био синтеза полисахарида — другой. Повышенные концентрации азо та в среде, как правило, отрицательно сказываются на синтезе полисахаридов. Качественный состав гликанов, по-видимому, не зависит от источников азота. Другие компоненты среды. На биосинтез полисахаридов мо гут влиять и другие необходимые для роста микроорганизмов компоненты среды. Большое значение для образования гликанов имеет фосфор. Повышенное содержание фосфора в среде тормо зит синтез многих полисахаридов. Но увеличение концентрации фосфора до определенных пределов способствует накоплению левана G. oxydans. Важное значение имеют различные ионы, необходимые для поглощения субстрата или в качестве кофакторов биосинтеза полисахаридов. Катионы железа оказывают положительное дей ствие на продукцию полисахаридов Pseudomonasaeruginosaи Methytomonasmucosa, на выделение в среду левансахаразы G. oxydans. Ионы марганца необходимы для образования глю-кана Rhyzobiamjaponicutnи маинана Sacch. cerevisiae. Магний способствует синтезу разветвленных декстранов L. mesenteroides. Концентрация ионов кальция в среде определяет соотношение маннуроновой и гулуроновой кислот в экзополисахариде A. vine-landii. Образование гликанов, как правило, не требует дополни тельных количеств витаминов сверх необходимого для нормаль ного роста продуцента. Стимуляция биосинтеза полисахаридов витаминными добавками наблюдается только в отдельных слу чаях. Физико-химические факторы среды Кислотность среды. Обычно существуют определенные грани цы рН, допускающие рост микроорганизмов и синтез полисаха ридов; эти интервалы у разных микроорганизмов неодинаковы. У ряда представителей границы, а иногда и оптимальные значения рН для роста и образования гликанов совпадают, но в отдельных случаях они различаются. У некоторых микроорга низмов количество полисахаридов не меняется в зависимости от рН среды. Чаще же изменение кислотности среды влияет на выход гликанов, на количественное соотношение полисахаридов, если их образуется несколько (аубазидан и пуллулан в культуре A. pullulans), на их молекулярную массу. Например, увеличение рН среды от 5,0 до 8,0 способствует образованию высокомоле кулярных фракций декстранов разными штаммами стрептокок ков, но мономерный состав гликанов при изменении рН среды, по-видимому, не меняется. Аэрация и температура. Влияние аэрации и температуры на биосинтез полисахаридов очень разнообразно. Режимы аэра ции и температуры, благоприятные для образования того или иного гликана, могут сильно различаться. Большинство микроорганизмов, образующих экзогликаны, — аэробы или факультативные анаэробы, поэтому в условиях хорошей аэрации выход зкзогликанов в культурах таких микро организмов выше. Однако избыточное аэрирование может угне тать их биосинтез вследствие быстрого окисления углеродного субстрата. В отношении действия температуры также наблюдается опре деленная закономерность. Максимальное образование гликанов часто происходит при температуре ниже оптимальной для роста микроорганизмов. Так, количество декстрана в культуре L. те-senieroidesувеличивается с уменьшением температуры с 30 до 10 °С. Но в некоторых случаях температурные оптимумы роста бактерий и образования ими экзогликанов совпадают. Изменение температуры культивирования микроорганизмов иногда приводит к синтезу полисахаридов с измененными свой ствами. Например, известен штамм стрептококков, синтезирую щий при 22 °С внеклеточный полисахарид, отличающийся более высоким содержанием глюкозамина и иными иммунохимическими свойствами, чем гликан, образуемый при 37 °С. Получение экзополисахаридов имеет преимущества перед получением внутриклеточных, так как экзогликаны образуются, как правило, в значительно большем количестве, легче отде ляются от биомассы и очищаются от примесей. Однако при производстве экзогликанов имеются свои технологические труд ности. Накопление полисахарида в среде приводит к ограниче нию доступа кислорода к клеткам. У аэробных микроорганизмов это снижает энергетический баланс и тормозит синтез полисаха рида. Повышенная вязкость среды делает невозможным отде ление полисахарида от клеток продуцента из нативной культу-ральной жидкости. Ее приходится разбавлять в десятки раз, а после удаления клеток концентрировать до первоначального или меньшего объема. Решение этих проблем связано с допол нительными затратами. Приведем основные этапы производства наиболее широко применяемых сейчас полисахаридов—декстрана и ксантана. Плазмозаменители из декстраиов выпускают под названиями: клинический декстран, полиглюкин, синкол, макродекс, плазмо-декс, хемодекс и др. Для получения декстрэнов используют штаммы Leucanostocmessnteroldes. Ферментацию ведут на среде с 10—30 % сахарозы, декстраном — «затравкой», дрожжевым экстрактом, минеральными солями. Создают условия, способст вующие синтезу той формы декстрана, которая используется в качестве плазмозаменителя: линейного глюкана, имеющего более 90% а-1,6-связей, с молекулярной массой 60—80 тыс. Для этого ограничивают содержание в среде магния, стимули рующего синтез разветвленных декстранов, вносят «затравку» в виде декстрана, имеющего молекулярную массу 20—30 тыс. Такой акцептор обеспечивает преимущественное образование не обходимого полимера. Наивысшей биологической активностью обладают декстраны, содержащие менее 70% ц-1,6-гликозидных связей, т. е. более разветвленные. Синтезу биологически активных декстранов спо собствует, кроме магния, замена сахарозы мелассой. Опти мальное значение рН для роста продуцента лежит в пределах 6,5—8,0, а для накопления декстрансахаразы — около 7,0. Обычно значение рН среды задают в интервале 7,0—8,0. Бактерии расщепляют сахарозу на глюкозу и фруктозу. Фруктоза сбраживается по типу гетероферментативного молочно кислого брожения с образованием молочной и уксусной кислот, маннита и СО2. Глюкоза полимеризуется в декстран. Процесс идет быстро и продукт можно выделить уже через 24 ч. 4 Технология получения декстрана 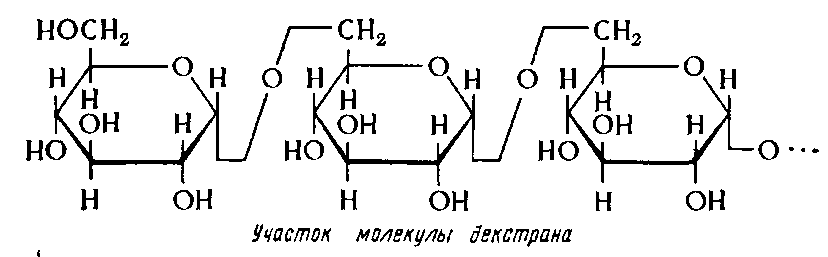 Декстран – разветвленный полисахарид бактериального происхождения, содержащий остатки глюкозы. Приготовленные на его основе гели выпускаются различными зарубежными фирмами и широко используются в различного рода работах, Некоторые из них известны под названием "сефадекс" (Швеция) и "молселект" (Венгрия). Гели, приготовленные на основе декстрана, отличаются высокой стойкостью по отношению к различным химическим веществам, что делает их весьма широко используемыми в различного рода исследованиях и на производстве. К группе декстранов может быть отнесен и крахмал, представляющий собой смесь полисахаридов, основным компонентом которой является амилоза и амилопектин. Посредством определенных химических обработок из крахмала получен новый носитель – губчатый крахмал, обладающий повышенной устойчивостью к ферментам, гидролизующим полисахариды. Продуцентами декстранов являются штаммы Leuconostac mesenteroides, растущие на средах с высоким содержанием сахарозы (10–30 %), дестраном-«затравкой», дрожжевым экстрактом и минеральными солями. В зависимости от состава минеральных солей и той или иной природы «затравки» синтезируются высокомолекулярные (60–80 тыс.) линейные или имеющие низкую молекулярную массу (20–30 тыс.) разветвленные декстраны. Последние обладают наибольшей биологической активностью. Из декстранов выпускают плазмозаменители (клинический декстран, по-лиглюкин, плазмодекс, хемодекс и др.). Типичный пример ферментации – глубинная периодическая культура, реализуемая на первом этапе с целью образования биомассы продуцента при избытке сахаров и рН 6.5–8.0. Синтез декстрансахаразы, ведущий к образованию гликанов, наиболее интенсивен при рН около 7.0. Помимо ионов магния синтез декстранов стимулируется при замене сахарозы мелассой. Бактерии расщепляют сахарозу с образованием глюкозы и фруктозы. Последняя сбраживается по гетероферментному пути с образованием молочной и уксусной кислот, маннита и углекислоты. Глюкоза быстро полимеризуется в декстран. Процесс завершается через 24 ч. Выделение декстрана из культуры проводят метанолом, для последующей очистки –многократно растворяют в воде, переосаждают метанолом и фракциони-руют. Декстрансахараза является экзоферментом, и ее концентрация в культуральной среде значительна. Поэтому возможен процесс получения полисахарида на основе растворимого фермента. Культуральная жидкость с декстрансахаразой при рН около 5.0 и 15°С способна около месяца про- являть высокую ферментативную активность. Реализован процесс на основе культуральной среды с ферментом, содержащей сахарозу и декстран- «затравку», – процесс полимеризации завершается в течении 8 ч. Этот способ значительно упрощает процедуру ферментации и стадию выделения и очистки декстрана и позволяет в контролируемых условиях получать продукт заданной молекулярной массы. Перспективы имеет также процесс на основе иммобилизованной декстрансахаразы. В середине 90-х гг. начат выпуск коньюгатов модифицированного декстрана с ферментом стрептокиназой. Препарат представляет собой пролонгированную декстраном форму стрептокиназы. Деполимеризованный декстран, молекулярная масса которо го близка к молекулярной массе альбумина сыворотки крови, используют в медицине как заменитель сыворотки крови. Для этого готовят 6%-ный раствор декстрана в физиологической жидкости, который по коллоидным и осмотическим свойствам близок к плазме крови. Этот раствор называют полигликином. Препарат полиглюкин широко используют в хирургии. Все станции скорой помощи обеспечены этим препаратом. При боль ших потерях крови вместо нее в кровеносную систему вводят полигликин. В опытах с собаками доказано, что даже при потере крови на 2/3 с помощью полигликина можно спасти жизнь. При восстановлении крови декстран постепенно используется на энергетические потребности. Надо отметить, что белки в такой ситуации не используются, так как организм очень резко реаги рует на присутствие чужеродных белков. Полигликин можно хранить в течение длительного времени. Ввести его в организм гораздо легче, чем переливать кровь. Однако этот препарат не может полностью заменить переливание крови, так как искусственно созданная система не в состоянии выполнить функцию переноса кислорода в организме. Для получения декстрана используют культуры Leuconostoc и L. dextranicum. Это шарообразные бактерии диаметром 1-1,5 мкм. В культуре они образуют переплетен ные цепочки. На твердой, содержащей сахарозу среде они обра зуют колонии, покрытые толстым слоем слизи. Главное исходное вещество при производстве полиглюкина сахароза. Ферментацию ведут методом глубинного культивиро вания. Концентрация сахарозы в среде должна быть высокой (око ло 20%), содержание азота — низким Для процесса полимеризации глюкозы присутствие кислорода воздуха не обязательно, но пере мешивание среды воздушным по током ускоряет реакцию. Надо отметить, что в процессе биосин теза декстрана большое значение имеет количество магния в среде. В зависимости от этого меняется строение молекулы декстрана, степень разветвленности, что в свою очередь воздействует на биологическую функцию препа рата в организме человека. Про цесс биосинтеза идет без участия фосфора. Биосинтез декстрана идет сле дующим образом: от молекулы сахарозы отщепляется молекула фруктозы, которая в своем метаболизме дает побочные продук ты, такие, как молочную и уксусную кислоты или многоатомный спирт маннит. Оставшиеся молекулы глюкозы под действием фермента декстрансахаразы полимеризуются. Оптимум pH дей ствия этого фермента от 5,2 до 5,6. Ферментация идет 20-24 ч 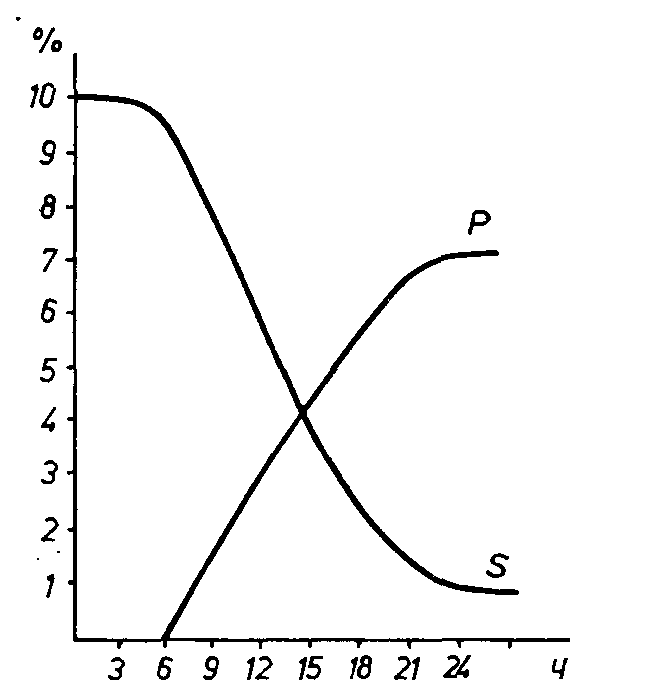 Рис. 1. Динамика биосинтеза декстрана: Р – декстран, S - сахароза Клинический декстран получают, обрабатывая нативный ра створ декстрана метиловым спиртом. Происходит деполимериза ция, и осаждается продукт с молекулярной массой 65000 - 80000. Деполимеризацию можно провести ферментативно, тер мически или с помощью ультразвука. Для получения более качественного клинического препарата процесс биосинтеза можно вести по методу введения ростков декстрана, которые не дают образовываться слишком крупным агрегатам молекул. Для этого в среду вносят акцепторы – зачатки (ростки) с молекулярной массой 20000 – 30000 (около 5% от количества сахарозы). В этом случае ферментация идет в среде, где содержится сахарозы 20%, зачатков декстрана – 2,9%, дрожжевого экстракта – 0,3%, двузамещенного фосфата натрия – 0,5%, хлорида калия – 0,1%, рН среды 6,5 – 8,0 5 Технология получения ксантана Ксантан, продуцируемый бактериями Xanthomonas campesrtis, обладает уникальными реологическими свойствами. В низких концентрациях он образует очень вязкие растворы и обладает псевдопластичностью; его растворы не изменяют свои реологические свойства при изменении температуры, рН, солености в широких пределах. Ксантан применяют в пищевой промышленности, при изготовлении гелевых дезодорантов, зубной пасты, при суспендировании сельскохозяйственных химикатов, используют при добыче нефти. Объемы производства ксантана – наиболее крупнотоннажны из всех других гликанов. Товарное название выпускаемого продукта (ксантан, келцан, келтрол). Получают ксантаны в условиях периодической глубинной культуры на средах, содержащих 1–5 % углеводов (кукурузный крахмал, сахар-сырец или меласса), а также органические соединения азота, двузамещенный фосфорнокислый калий, микроэлементы. Ферментация длится в течение 3 суток при 28°С и рН 6.5–7.2 в две фазы: на первой реализуется рост клеток и накопление биомассы, на второй при дефиците азота в среде происходит образование полисахарида. Осаждают полисахарид из культуральной жидкости метанолом, полученный осадок высушивают. 6 Технология получения альгината Данный полисахарид ранее выделяли из морской водоросли Laminaria. Альгинат обладает в определенных условиях прекрасными гелеобразующими, а также псевдопластическими свойствами в широком диапазоне рН и температур, и используется в кондитерской и фармацевтической промышленности. Установлено, что альгинат является лучшим носителем для иммобилизации ферментов и, особенно, целых клеток. Сравнительно недавно среди бактерий идентифицированы продуценты полисахарида, близкого альгинату (Pseudomonas aeruginosa, Azotobacter vinelandii). Процесс реализован в промышленности на средах с избытком углерода. Варьируя концентрацию фосфата в среде, можно влиять на молекулярную массу синтезируемого полимера, а при изменении концентрации кальция из- меняется соотношение моносахаридов, входящих в состав данного гликана, следовательно, и его свойства. Заключение В ходе курсовой работы дана характеристика биополимеров, а также полисахаридов на примере декстрана. Была рассмотрена характеристика микроорганизмов – продуцентов полисахаридов, условия культивирования, методы культивирования. В предложенной схеме производства декстрана дана динамика биосинтеза, его характеристика, применение, параметры роста, режимы культивирования. Задачей биотехнологии является совершенствование микробиологических процессов получения полисахаридов на основе улучшенных штаммов-продуцентов при расширении сырьевой базы за счет замены дорогостоящих сахаров более доступными субстратами, а также модификация физико-химических свойств самих гликанов. Список литературы: 1. Божков А.И. Биотехнология. Фундаментальные и промышленные аспекты. Учебное пособие для студентов высших учебных заведений. Харьков. - 2005. - 364с. 2. Волова Т. Г. Биотехнология / Т. Г. Волова. – Новосибирск: Изд-во Сибирского отделения Российской Академии наук, 1999. – 252 с. 3. Евтушенков А.Н., Фомичев Ю.К. Введение в биотехнологию: Курс лекций:/ А.Н.Евтушенков, Ю.К.Фомичев. - Мн.: БГУ, 2002. - 105 с. 4. Егорова Т.А., Клунова С.М., Живухина Е.А. Основы биотехнологии. - М.: Издательский центр «Академия», 2003. - 208 с. 5. Елинов Н.П. Основы биотехнологии. СПб "Наука", 1995. - 601 с Коростелева Н.И. Биотехнология: учебное пособие / Н.И. Коростелева, Т.В. Громова, И.Г. Жукова. Барнаул: Изд-во АГАУ, 2006. - 127 с. 6. Катлинский А.В. Курс лекций по биотехнологии. - М.: Московская медицинская академия им. И. М. Сеченова, 2005. - 152 с. 7. Красноштанова А.А., Крылов И.А., Бабусенко Е.С. — Основы биотехнологии. — М., Изд-во РХТУ им.Д.И.Менделеева, 2000. — 84 с. 8. Макаров С. В., Никифорова Т. Е., Козлов Н. А. Основы биотехнологии: Учеб. пособие / ГОУ ВПО Иван. гос. хим.-тех. ун-т. Иваново, 2005. - 48 с. 9. Сазыкин Ю.О., Орехов С.Н., Чакалева И.И.; Биотехнология: Учебное пособие / Под ред. А.В. Катлинского. – 2 – е изд., стер.- М.: Академия, 2007. – 254с. 10. Тихонов И.В., Рубан Е.А., Грязнева Т.А., и тд. Биотехнология: Учебник / Под ред. Е.С. Воронина.- СПб.: ГИОРД, 2008. - 703с. |