Дубровский В.И., Федорова В.Н. Биомеханика. Учебник для вузов
 Скачать 6.47 Mb. Скачать 6.47 Mb.
|

17.2. Биомеханика дыханияДыхание Обмен кислорода (О2) и углекислоты (СО2) между организмом и средой называется дыханием. Человеческий организм в процессе жизнедеятельности потребляет кислород (О2) и выделяет углекислоту (СО2). Здоровый мужчина среднего возраста и нормального сложения с массой тела 70 кг в условиях основного обмена потребляет за 1 мин. 250 мл О2 и выделяет около 200 мл углекислоты. При физической нагрузке потребление О2 и, соответственно, выделение СО2 увеличивается в несколько раз. При этом повышение тканевого обмена обеспечивается не только пропорциональным увеличением потребления О2, возрастает также утилизация О2, в результате чего происходит более полное восстановление оксигемоглобина в тканях. Обеспечение организма нужным количеством О2 и выведением СО2 возможно лишь при условии нормального течения и координированного изменения ряда последовательных актов. У человека дыхание осуществляется благодаря ряду последовательных процессов: 1) обмен газов между средой и легкими, что обычно обозначают как «легочную вентиляцию»; 2) обмен газов между альвеолами легких и кровью (легочное дыхание); 3) обмен газов между кровью и тканями. Наконец, газы переходят внутри ткани к местам потребления (для О2) и от мест образования (для СО2) (клеточное дыхание). Движение газов в дыхательной системе и между средой и тканями происходит в результате разницы давлений. Пониженное давление О2 в ткани заставляет газ двигаться к ней. Для СО2 градиент давления направлен в обратную сторону, и СО2 переходит в окружающую среду. Известно, что давление водяных паров в организме выше, чем в окружающей среде, и, таким образом, при дыхании организм теряет воду. Дыхательная система (рис. 17.13) состоит из тканей и органов, обеспечивающих легочную вентиляцию и легочное дыхание (воздушные пути, легкие и элементы костно-мышечной системы). К воздухоносным путям относятся: нос, полость носа, носоглотка, гортань, трахея, бронхи и бронхиолы. Легкие состоят из бронхиол и альвеолярных мешочков, а также из артерий, капилляров и вен легочного круга кровообращения. К элементам костно-мышечной системы, связанным с дыханием, относятся ребра, межреберные мышцы, диафрагма и вспомогательные дыхательные мышцы. Легкие Легкие представляют собой важнейшую структуру, осуществляющую физиологическую связь организма с окружающей средой: общая площадь их поверхности примерно в 30 раз больше, чем площадь кожи. 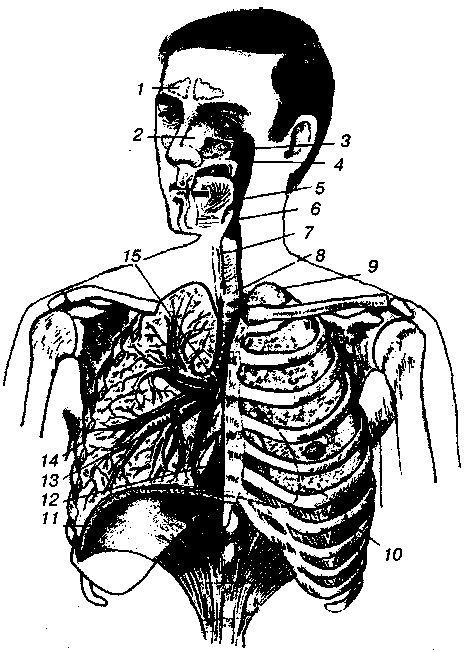  Рис. 17.13. Дыхательная система: 1 — лобная пазуха, 2 — решетчатая кость, 3 — гайморова пазуха, 4 — носоглотка, 5 — ротовая часть глотки, 6 — гортанная часть глотки, 7 — гортань, 8 — трахея, 9 — верхушка легкого, 10 — межреберные мышцы, И—диафрагма, 12 — сердце, 73 — ветви нижнедолевого бронха, 14 — ребра, /5 — правый бронх, 16 — альвеолы В целом легкие имеют вид губчатых, пористых конусовидных образований, лежащих в обеих половинах грудной полости. Наименьший структурный элемент легкого — долька состоит из конечной бронхиолы, ведущей в легочную бронхиолу и альвеолярный мешок. Стенки легочной бронхиолы и альвеолярного мешка образуют углубления — альвеолы. Стенки альвеол состоят из одного слоя эпителиальных клеток типа I и окружены легочными капиллярами. Принято считать, что общая поверхность альвеол, через которую осуществляется газообмен, экспоненциально зависит от веса тела. С возрастом отмечается уменьшение площади поверхности альвеол. Плевра Каждое легкое окружено мешком, образованным серозной оболочкой — плеврой (рис. 17.14). Наружный (париентальный) листок плевры примыкает к внутренней поверхности грудной стенки и диафрагме, внутренний (висцеральный) покрывает легкое. Щель между листками называется плевральной полостью. При движении грудной клетки внутренний листок обычно легко скользит по наружному. Давление в плевральной полости всегда меньше атмосферного (отрицательное). В условиях покоя внутриплевральное давление у человека в среднем на 4,5 торр ниже атмосферного (—4,5 торр). 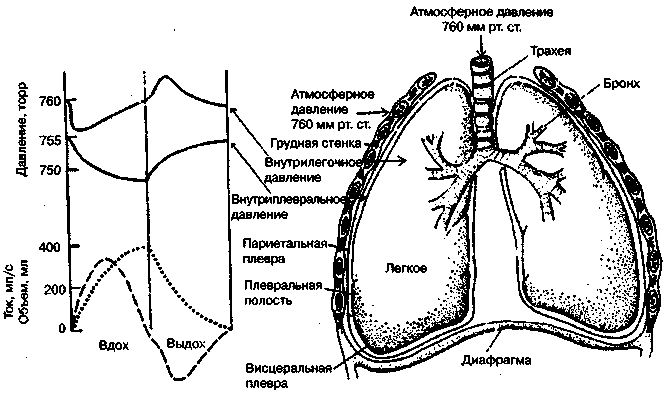 Рис. 17.14. Изменения внутрилегочного и внутриплеврального давления на протяжении цикла вдох—выдох. Показаны также изменения скорости тока воздуха и объема легких Грудная полость Грудная полость ограничена сзади первыми десятью грудными позвонками (см. рис. 17.13), последние два грудных позвонка функционально относятся к брюшной полости и не принимают активного участия в дыхании. Переднюю стенку грудной клетки образует грудина. Боковая стенка грудной клетки образована ребрами и реберными хрящами. Ребра лежат парами по обе стороны позвоночника. Каждое ребро наклонено вниз от уровня своего сочленения с позвонком и прикреплено к грудине ниже (см. рис. 17.16). Пространства между ребрами называется межреберным. Дыхательные мышцы Дыхательные мышцы — это те мышцы, сокращения которых изменяют объем грудной клетки. Мышцы, направляющиеся от головы, шеи, рук и некоторых верхних грудных и нижних шейных позвонков, а также наружные межреберные мышцы, соединяющие ребро с ребром, приподнимают ребра и увеличивают объем грудной клетки. Диафрагма — мышечно-сухожильная пластина, прикрепленная к позвонкам, ребрам и грудине, — отделяет грудную полость от брюшной (рис. 17.15). Это главная мышца, участвующая в нормальном вдохе. При усиленном вдохе сокращаются дополнительные группы мышц. При усиленном выдохе действуют мышцы, прикрепленные между ребрами (внутренние межреберные мышцы) к ребрам и нижним грудным и верхним поясничным позвонкам, а также мышцы брюшной полости; они опускают ребра и прижимают брюшные органы к расслабившейся диафрагме, уменьшая таким образом емкость грудной клетки. 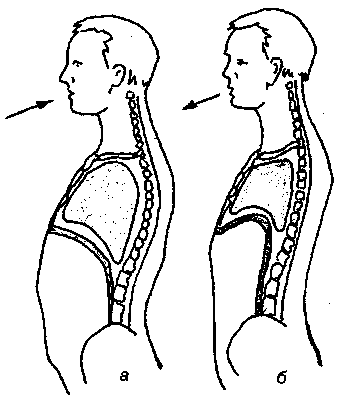 Рис. 17.15. Движения диафрагмы: а — положение диафрагмы при вдохе, б — при выдохе. Положение диафрагмы (вид сбоку) во время дыхания (в) 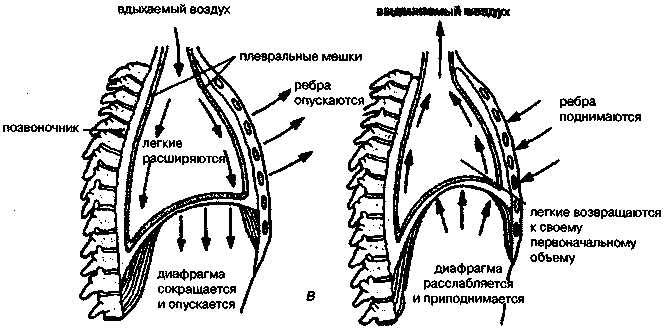 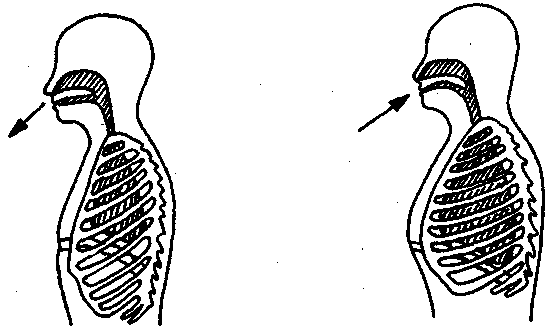 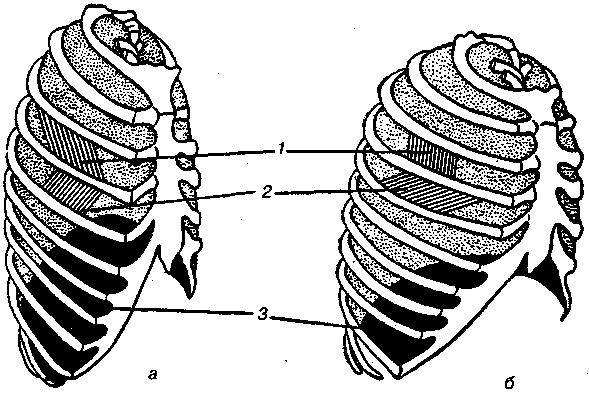 Рис. 17.16. Схема положения грудной клетки и диафрагмы при выдохе (а) и вдохе (б): 1 — наружные межреберные мышцы, 2 — внутренние межреберные мышцы, 3 — диафрагма Дыхательные движения осуществляются за счет дыхательной мускулатуры. Расслабление всех связанных с дыханием мышц придает грудной клетке положение пассивного выдоха. Соответствующая мышечная активность может перевести это положение во вдох или же усилить выдох. Механизм, вдоха Акт вдоха (инспирация) совершается вследствие увеличения объема грудной полости в трех направлениях — вертикальном, сагиттальном и фронтальном. Это происходит вследствие поднятия ребер и опускания диафрагмы (рис. 17.16). В состоянии выдоха ребра опущены вниз; а в состоянии вдоха — принимают более горизонтальное положение, поднимаясь кверху; при этом нижний конец грудины отходит вперед. Благодаря движению ребер при вдохе сечение грудной клетки становится больше и в поперечном, и в продольном направлениях (см. рис. 17.16). Ребра представляют собой рычаги второго рода с точкой вращения в их сочленениях с позвоночником (рис. 17.17, а, с). Наружные межреберные мышцы при сокращении должны были бы сближать ребра, но так как момент силы у нижнего прикрепления мышц (g) больше, чем у верхнего (b) вследствие большой длины рычага (c—g), то при сокращении мышц ребра поднимаются. 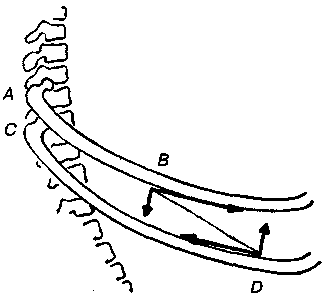 Рис. 17.17. Механика движений ребер при вдохе Во время вдоха диафрагма сокращается, в результате чего ее купол становится более плоским и опускается (см. рис. 17.16). В зависимости от возраста, пола, вида деятельности дыхание совершается преимущественно или за счет межреберных мышц — реберный, или грудной тип дыхания, или за счет диафрагмы — диафрагмальный, или брюшной тип дыхания. Есть и смешанный тип, при котором в дыхании участвуют нижние отделы грудной клетки и верхняя часть живота, он встречается у пожилых людей, а также при ригидности грудной клетки и снижении эластичности легочной ткани (эмфизема легких, пневмосклероз и др.). Тип дыхания не является строго постоянным и может меняться в зависимости от исходного положения, телосложения, пола, вида деятельности и состояния пациента. Так, при переносе на спине тяжелого груза грудная клетка фиксируется мышцами туловища и межреберий неподвижно вместе с позвоночником; дыхание же совершается исключительно за счет движений диафрагмы. У беременных женщин смещение диафрагмы вниз затруднено и поэтому преобладает реберный тип дыхания. При усиленном дыхании, например, у спортсменов, в акте вдоха участвует ряд дополнительных, или вспомогательных дыхательных мышц. При вдохе объем грудной клетки и находящихся в ней легких увеличивается; при этом давление в них понижается и воздух через воздухоносные пути входит в легочные альвеолы. Механизм выдоха Во время вдоха дыхательные мышцы человека преодолевают ряд сил: 1) тяжесть приподнимаемых кверху ребер; 2) эластическое сопротивление реберных хрящей; 3) сопротивление стенок живота и брюшных внутренностей, отдавливаемых книзу опускающимся куполом диафрагмы. Когда вдох окончен и дыхательные мышцы расслабляются, под влиянием указанных сил ребра опускаются и купол диафрагмы приподнимается. Объем грудной клетки вследствие этого уменьшается. Таким образом, акт выдоха (экспирация) происходит обычно пассивно, без участия мышц. При форсированном выдохе к перечисленным силам, уменьшающим объем грудной клетки, присоединяется сокращение внутренних межреберных мышц, задних нижних зубчатых мышц и мышц живота. При сокращении внутренних межреберных мышц, ребра опускаются. Мышцы живота при их сокращении оттесняют органы брюшной полости и купол диафрагмы кверху. При выдохе объем грудной клетки, а, следовательно, и легких, уменьшается, давление в альвеолах повышается и воздух выходит из легких наружу. У здорового человека дыхание в спокойном состоянии ритмичное, и число дыхательных движений составляет 14—18 в мин., а у спортсменов — 8—12. Дыхание может быть учащенным и редким. Учащение дыхания наблюдается после физической нагрузки (в процессе тренировки), при нервном возбуждении и др. Урежение дыхательных движений наблюдается при заболеваниях, которые угнетают функции дыхательного центра или при анатомических изменениях в бронхах (сужение, сдавливание и т. п.). У здорового человека дыхание ритмичное, глубокое. Но встречается и нарушение ритмичности дыхания, которое, как правило, является результатом нарушения координационной способности дыхательного центра, характеризующееся тем, что нарушается гармоническая, слаженная работа отдельных групп дыхательных мышц. В этой связи наступает более быстрая утомляемость дыхательной мускулатуры, что приводит к нарушению снабжения мышц кислородом и утомляемости пациента. Ритм дыхания может нарушаться при беге по пересеченной местности (кросс), у лыжников-гонщиков и в других видах спорта, а также при тестировании спортсменов с явлениями перетренированности (например, при выполнении пробы «степ-тест», или «бег на месте»). Механика дыхательных движений Перемещение воздуха в легкие и из них требует совершения работы. Для того, чтобы воздух вошел в легкие, должны быть преодолены силы трех типов, а именно: 1) эластическое сопротивление; 2) сопротивление воздушного потока в трахеобронхиальном дереве и 3) сопротивление неэластичных тканей, например, ребер. Расширение легких обусловлено увеличением объема грудной клетки. Если давление снаружи становится выше атмосферного, из легких выходит лишь небольшое количество воздуха, так как мелкие воздухоносные пути спадаются, задерживая его в альвеолах. С возрастом, а также при некоторых легочных заболеваниях такое закрытие дыхательных путей происходит при большем объеме легких. Крутизна кривой «давление—объем», т. е. изменение объема на единицу изменения давления, называется растяжимостью. В физиологических условиях (если растягивающее давление составляет от —2 до —10 см вод. ст.) легкие обладают удивительной растяжимостью. У человека она достигает примерно 200 мл/см вод. ст., однако при более высоких давлениях уменьшается. Этому соответствует более пологий участок кривой «давление—объем». Растяжимость легких несколько снижается при повышенном давлении в легочных венах и переполнении легких кровью. При альвеолярном отеке она уменьшается в результате неспособности некоторых альвеол раздуваться. Заболевания, сопровождающиеся фиброзом легких, воспалительными процессами, также приводят к уменьшению их растяжимости. Это связано с изменениями эластических тканей. В стенках альвеол, а также вокруг сосудов и бронхов проходят волокна эластина и коллагена. По определению, растяжимость легких равна изменению их объема на единицу изменения давления. Для ее оценки необходимо измерить внутриплевральное давление. При этом регистрируют давление в пищеводе: обследуемый заглатывает катетер с маленьким баллончиком на конце. Растяжимость легких можно измерить очень просто: обследуемого просят сделать максимально глубокий вдох, а затем выдыхать воздух в спирометр порциями, скажем, по 500 мл. При этом определяют давление в пищеводе. Затем строят график «давление—объем», сходный с кривой на рис. 17.18. Этот метод позволяет получить наибольшую информацию об упругости легких. Растяжимость легких можно также измерить при спокойном дыхании (рис. 17,19). Этот способ основан на том, что в отсутствие потока воздуха (в конце вдоха и выдоха) внутриплевральное давление отражает только эластическую тягу легких и не зависит от сил, возникающих при движении воздушной струи. Таким образом, растяжимость будет равна отношению разности легочных объемов в конце вдоха и выдоха к разности внутриплевральных давлений в эти же моменты. На вентиляцию легких влияют: частичное перекрытие (закупорка) воздухоносных путей (мокрота, слизь и др.) и тогда заполнение воздухоносных путей (участков легких) будет происходить медленнее. С увеличением частоты дыхания объем воздуха, поступающего на такой участок, становится все меньше и меньше. Упругостью обладают не только легкие, но и грудная клетка. В норме грудная клетка стянута, а легкие растянуты и действующие в них упругие силы уравновешивают друг друга. В эксперименте показано, что при объеме, равном функциональной остаточной емкости (ФОБ), давление релаксации отрицательно. Это означает, что грудная клетка стремится расшириться. Лишь в том случае, когда объем достигает примерно 75% жизненной емкости легких (ЖЕЛ), давление релаксации становится равным атмосферному, т. е. грудная клетка приходит в состояние равновесия. При любом объеме давление релаксации легких и грудной клетки равно сумме их давлений релаксации, измеренных по отдельности. Поскольку давление (при данном объеме) обратно пропорционально растяжимости, общую растяжимость легких и грудной клетки можно вычислить по формуле где Со — общая растяжимость легких и грудной клетки, Сл — растяжимость легких, Сгк — растяжимость грудной клетки. 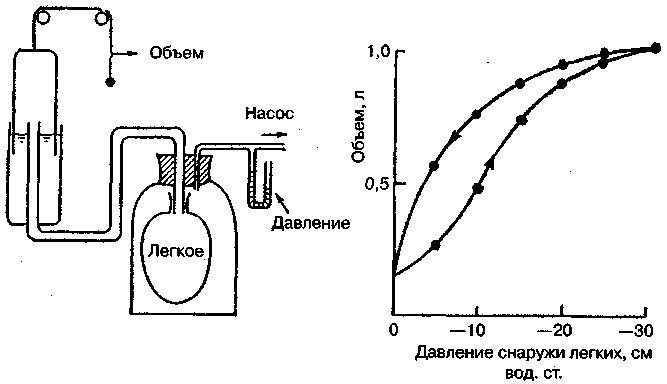 Рис. 17.18. Построение кривой зависимости объема от давления для изолированного легкого. Для этого вокруг легкого на несколько секунд создается определенное давление и измеряется легочный объем. Полученная кривая нелинейна: при высоких давлениях она становится более пологой. Видно, что кривые для раздувания и спадения легких неодинаковы; это явление называется гистерезисом Еще один важнейший фактор, во многом обусловливающий особенности кривых «давление—объем» для легких, — это поверхностное натяжение жидкости, выстилающей стенки альвеол. Поверхностным натяжением называется сила (измеряемая обычно в динах), действующая в поперечном направлении на воображаемый отрезок длиной 1 см на поверхности жидкости. Известно, что клетки, выстилающие стенки альвеол, вырабатывают секрет, значительно снижающий поверхностное натяжение альвеолярной жидкости. Влияние секрета (сурфактанта) на поверхностное натяжение, объясняется его низким поверхностным натяжением в альвеолах и отсюда увеличивается растяжимость легких и тем самым уменьшается совершаемая при вдохе работа; а также обеспечивается стабильность альвеол, их в легких около 300 млн, и все они имеют тенденцию к спадению (ателектазу), очаги которого часто образуются в легких при заболеваниях. При недостатке сурфактанта легкие становятся более «жесткими» (т. е. менее растяжимыми). Известно, что нижние отделы легких вентилируются лучше, чем верхние. Это, по-видимому, связано с тем, что в области оснований легких внутриплевральное давление менее отрицательно, чем в области верхушек.  Рис. 17.19. Изменение давления во время дыхания. При отсутствии аэродинамического сопротивления альвеолярное давление было бы всегда равно нулю, а внутриплевральное изменялось бы в соответствии с пунктирной кривой АБВ (то есть под действием лишь эластической тяги легких). Поскольку же дыхательные пути (и ткани) обладают вязким сопротивлением, внутриплевральное давление изменяется по сплошной кривой. «Вкладу» этого сопротивления соответствует заштрихованный участок Сопротивление воздухоносных путей Воздух проходит через трубку (рис. 17.20), если между ее концами существует перепад давлений. От его величины зависят скорость и особенности воздушного потока. При низких скоростях линии течения могут быть параллельны стенкам трубки (А). Это так называемый ламинарный режим. По мере возрастания скорости потока он становится все менее однородным, особенно в местах ветвления трубки, где разделение воздушных струй может происходить с образованием местных завихрений (Б). Наконец, при очень высоких скоростях линии течения полностью теряют упорядочность, и поток называется в этом случае турбулентным (В). Уравнение, связывающее давление и расход (т. е. объемную скорость) при ламинарном потоке было впервые выведено французским врачом Пуазейлем. Для прямых трубок с круглым сечением оно записывается следующим образом: j*# 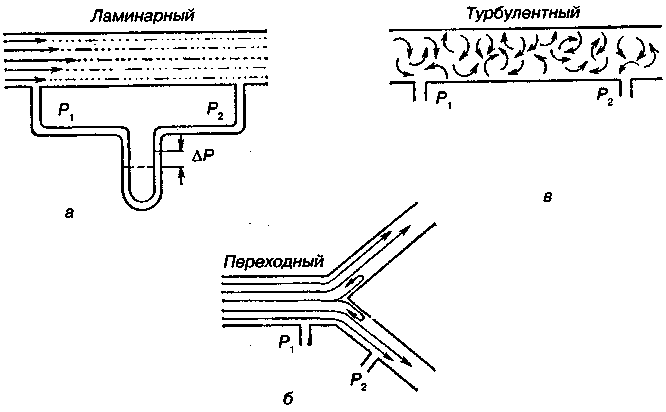 Рис. 17.20. Типы воздушного потока в трубках. а — ламинарный поток; б — переходный тип (с завихрениями в области ветвлений); в — турбулентный поток. Аэродинамическое сопротивление равно отношению перепада давлений (Р1—Р2) к расходу где V— расход флюида, Р — давление, создающее поток (ΔР см. на рис. 17.20), r — радиус трубки, η — вязкость флюида, l — длина трубки. Из уравнения видно, что давление пропорционально расходу (Р = KV). Поскольку сопротивления потоку R равно давлению, деленному на расход, можно записать: Как видно, большую роль играет радиус трубки; когда он уменьшается вдвое, сопротивление потоку увеличивается в 16 раз. Важно также, что на взаимоотношение между давлением и расходом влияет вязкость, а не плотность флюида. Одна из особенностей полностью развитого ламинарного потока заключается в том, что частицы газа в центре трубки передвигаются со скоростью, в два раза превышающей среднюю (см. рис. 17.20). Особенности турбулентного потока совершенно иные. Давление в этом случае пропорционально уже не расходу флюида, а примерно квадрату расхода (Р = KV2). Вязкость при таком режиме не играет существенной роли, а увеличение плотности флюида при данном расходе повышает перепад давлений. Будет поток ламинарным или турбулентным, в значительной степени зависит от так называемого числа Рейнольдса (Re), получаемого по уравнению: где d — плотность флюида, v — средняя линейная скорость, r— радиус трубки, η — вязкость флюида. В прямых гладких трубках турбулентность возможна при числе Рейнольдса больше 2000. Применить все эти закономерности к такой сложной системе трубок, как бронхиальное дерево — со всеми его разветвлениями, изменениями диаметра и неровными стенками —трудно. Практически особенности потока очень сильно зависят от «входных» характеристик трубки. Если у какой-либо развилки возникает завихрение, воздушная струя как бы «тянет» его за собой, и оно исчезает лишь на определенном расстоянии от места зарождения. Поскольку же бронхиальное дерево постоянно ветвится, можно полагать, что истинный ламинарный поток (см. рис. 17.20) возникает лишь в самых мелких воздухоносных путях, где число Рейнольдса очень мало (в конечных бронхиолах оно может составлять около 1). На остальных участках течение носит переходный характер (Б). Турбулентный поток может наблюдаться в трахее, особенно при физической нагрузке, когда скорость воздуха возрастает. В целом для расчета «перепада» давления в бронхиальном дереве следует использовать как первую, так и вторую степень расхода воздуха: P=K1V+K2V2. Сопротивление воздухоносных путей можно рассчитать, разделив разность давления в ротовой полости и альвеолах на расход воздуха (см. рис. 17.20). В ротовой полости давление легко измеряется с помощью манометра, а в альвеолах его можно оценить с помощью общего плетизмографа. Сопротивление воздухоносных путей равно отношению разности давлений между альвеолами и ротовой полостью к расходу воздуха (см. рис. 17.20). Его можно измерить методом общей плетизмографии (рис. 17.21). Перед тем, как обследуемый делает вдох (А), давление в плетизмографической камере равно атмосферному. Во время вдоха давление в альвеолах снижается, а объем альвеолярного воздуха увеличивается на величину ΔV. При этом воздух в камере снижается и по изменению его давления можно рассчитать ΔV (см. рис. 17.22). 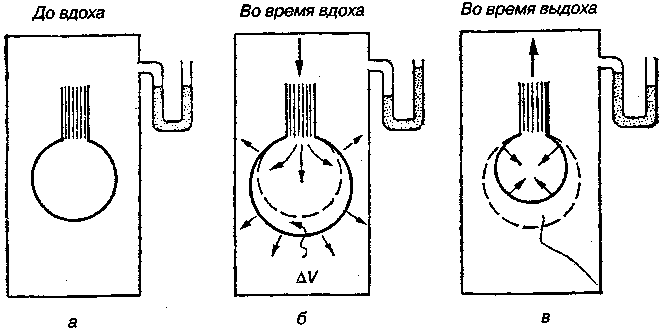 Рис. 17.21. Измерение сопротивления воздухоносных путей с помощью общей плетизмографии. Во время вдоха альвеолярный воздух расширяется, и давление в камере увеличивается. По этому увеличению можно рассчитать внутриальвеолярное давление. Разделив разницу между давлением в альвеолах и полости рта на расход воздуха, можно получить сопротивление воздухоносных путей (по J. Comrol, 1965) Р1 V1= Р2(V1- ΔV), где Р1и Р2— давление в камере соответственно до попытки вдохнуть и во время нее, V1— объем камеры до этой попытки, а ΔV — изменение объема камеры (или легких). Отсюда можно рассчитать ΔV. Если объем легких известен, можно перейти от ΔV к внутриальвеолярному давлению, используя закон Бойля-Мариотта (Р3V2 = P4(V2+ ΔV ), где Р3и Р4 — давление в полости рта соответственно до попытки вдохнуть и во время нее, a V2— ФОЕ, которая и рассчитывается по этой формуле). Одновременно измеряется расход воздуха, что дает возможность рассчитывать сопротивление воздухоносных путей. Такие же измерения проводятся при выдохе. Способ определения объема легких приведен на рис. 17.22. Сопротивление воздухоносных путей можно рассчитать и при спокойном дыхании, измерив внутриплевральное давление с помощью введенного в пищевод катетера (см. рис. 17.19). Однако при этом результаты будут включать также сопротивление тканей. Внутриплевральное давление определяется с одной стороны силами, противодействующими эластической тяге легких, а с другой — силами, преодолевающими сопротивление воздухоносных путей и тканей. 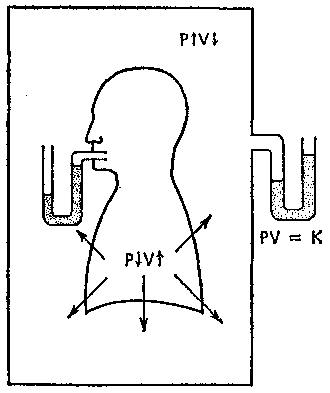 Рис. 17.22. Измерение ФОЕ с помощью общей плетизмографии. Когда обследуемый пытается сделать вдох при перекрытых воздухоносных путях, объем его легких несколько увеличивается, давление в дыхательных путях снижается, а давление в камере повышается. Отсюда, используя закон Бойля—Мариотта, можно рассчитать объем легких При движении легких и грудной клетки необходимо прикладывать некоторое давление для преодоления вязких сил, действующих в тканях при их деформации. Именно наличием таких сил частично объясняется заштрихованная область кривой на рис. 17.19. Однако у молодых здоровых людей сопротивление тканей составляет лишь около 20% общего (т. е. суммы сопротивления тканей и воздухоносных путей), хотя при некоторых заболеваниях оно может увеличиваться. Для того, чтобы при дыхании происходили движения легких и грудной клетки, необходимо затрачивать работу. В данном случае ее удобнее всего измерять произведением давления на объем. Работу, затрачиваемую на движение легких можно оценить по кривой «давление—объем» (рис. 17.23). При вдохе внутриплевральное давление изменяется в соответствии с кривой АБВ и на движение легких затрачивается работа, соответствующая площади ОАБВГО. Трапеция ОАДВГО отражает работу, необходимую для преодоления упругих сил, а заштрихованный участок АБВДА — работу, затраченную на преодоление вязкого сопротивления воздухоносных путей и тканей (см. рис. 17.19). Чем выше сопротивление воздухоносных путей или расход воздуха при вдохе, тем более отрицательным будет внутриплевральное давление, тем больше сместится вправо (в сторону отрицательных величин) точка Б по сравнению с точкой Д и тем больше будет площадь заштрихованного участка. 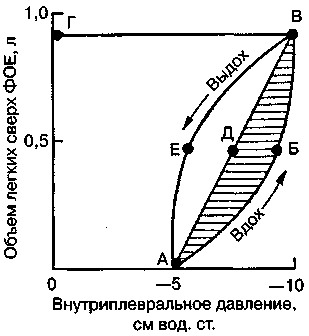 Рис. 17.23. Кривая «давление—объем» для легких. Работа, необходимая для преодоления упругих сил при вдохе соответствует трапеции ОАДВГО, а работа по преодолению вязких сил — заштрихованному участку АБВДА Работе, необходимой для преодоления сопротивления воздухоносных путей (и тканей) при выдохе, соответствует участок АДВЕА. В нормальных условиях он «вписан» в трапецию ОАДВГО, т. е. работа по преодолению вязких сил может быть совершена за счет энергии, запасенной в упругих структурах и высвобождающейся при пассивном выдохе. Разница между площадями АДВЕА и ОАДВГО соответствует энергии, рассеивающейся в виде тепла. Чем выше частота дыхания и расход воздуха, тем больше площадь участка АБВДА (т. е. работа по преодолению вязких сил). С другой стороны, чем больше дыхательный объем (ДО), тем больше площадь трапеции ОАДВГО (т. е. работа по преодолению упругих сил). Больные со сниженной растяжимостью легких (пневмосклероз, эмфизема и др.) как правило, дышат чаще и дыхание поверхностное; а при обструкции дыхательных путей — дыхание медленное. В обоих случаях это способствует уменьшению затрачиваемой работы. При выполнении тяжелой физической работы, при занятиях спортом, особенно циклическими видами спорта (академическая гребля, плавание, лыжные гонки, стайерский бег и др.) происходит увеличение затрачиваемой работы, и затраты увеличиваются, если спортсмен тренируется в неблагоприятных климатических условиях (среднегорье, зоны с жарким и влажным климатом и т. п.). Общую работу, затрачиваемую на движение легких и грудной клетки, измерить трудно, хотя некоторые ее оценки были получены при искусственной вентиляции в респираторе типа «искусственные легкие». Такую работу можно рассчитать так же, измеряя затраты кислорода на дыхание и учитывая коэффициент полезного действия (КПД): Полагают, что этот коэффициент составляет около 5—10%. Затраты кислорода на спокойное дыхание исключительно малы — менее 5% от общего потребления О2. При произвольной гипервентиляции они могут увеличиваться до 30%. У спортсменов во время физической работы (тренировки или соревнования) поглощение кислорода дыхательными мышцами увеличивается и тем самым дыхательная мускулатура является лимитирующим фактором в выполнении физической работы (нагрузки). Работа, необходимая для преодоления эластического сопротивления легких и грудной стенки, как полагают, не зависит от времени. Максимум работы производится тогда, когда дыхательный объем также максимален. Эту форму сопротивления можно вычислить, определив давление, необходимое для измерения объема легких и грудной клетки. Эта величина называется растяжимостью (С). где ΔV — изменение объема, а ΔР — изменение давления. Общую растяжимость легкого и грудной стенки можно определить, составив график, выражающий внутрилегочное давление, необходимое для поддержания в легком известного объема газа. Экспериментально это производится путем наполнения легких неким объемом, расслабления всех дыхательных мышц и измерения давления во рту (при закрытых ноздрях). Растяжимость легкого равна величине внутриплеврального давления и может быть определена таким же образом (рис. 17.24). 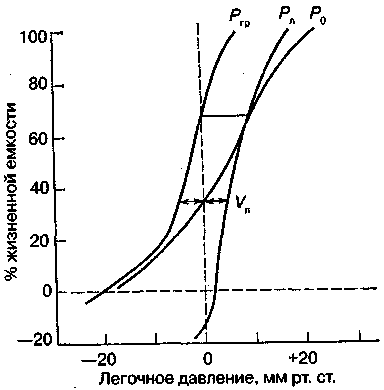 Рис. 17.24. Общее давление (Р0), создаваемое суммой эластических свойств грудной клетки (Ргр) и легких (Рл) на разных уровнях расширения груди (%ЖЕ). Наклон кривых соответствует растяжимости. Обратите внимание на то, что конечный дыхательный объем в покое (Vn) приходится на точку, где отрицательное (Ргр) равно положительному (Рл). При изменении эластических свойств легкого или грудной клетки (Vn) должно сместиться. Любой другой объем кроме Vn, требует напряжения мышц для создания нужной силы (Р0) Установлено, что от 3/4 до 7/8 общего эластического сопротивления создается поверхностным натяжением пленки жидкости, выстилающий внутреннюю поверхность альвеолы, а остальная часть — эластическими свойствами ткани. Чем выше поверхностное натяжение, тем больше нужно энергии для преодоления его сопротивления. Поверхностное натяжение снижается за счет сурфактанта. Как полагают, сурфактант стабилизирует легочные альвеолы, так что они не спадаются при выдохе. Показано, что сопротивление воздушному потоку создается главным образом в бронхах среднего размера (рис. 17.25). На основании уравнения Пуазейля следовало бы ожидать, что местом наибольшего сопротивления будут самые мелкие бронхиолы, но на самом деле это не так. Воздушные пути с диаметром меньше 2 мм создают менее 20% измеренного сопротивления воздушному потоку. Обилие мелких воздушных путей создает большое суммарное поперечное сечение для воздушного потока. Для очень малого объема легких описано явление «закрытого воздухоносного пути», т. е. обратимого спадения мелких бронхиол. В таких условиях некоторое количество энергии затрачивается при вдохе на открывание спавшихся бронхиол. Сопротивление воздушному потоку зависит от времени; оно наибольшее при частом дыхании и достигает максимума, даже если объем вдоха не максимален. 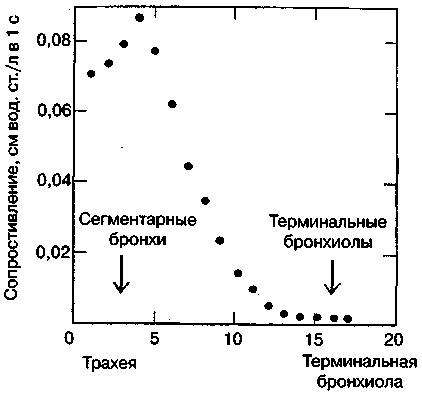 Рис. 17.25. Сопротивление в разных частях воздухоносных путей. Обратите внимание на то, что сопротивление выше всего в крупных бронхах, а самое низкое — в мелких бронхиолах Работа по перемещению грудной клетки и легкого против сопротивления неэластичных тканей тоже зависит от времени. У взрослых мужчин она составляет около 20% общего расхода энергии при дыхании. Общую работу, затрачиваемую на перемещение воздуха в легкое и из него, включая движение грудной клетки, можно вычислить по графику «давление—объем» (рис. 17.26): 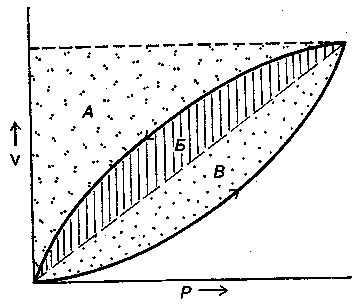 Рис. 17.26. Схема зависимости между давлением и объемом при одном дыхательном объеме (сплошная линия, направленная в сторону возрастающих значений, обозначает вдох; сплошная линия, направленная в сторону снижения объема, обозначает выдох). Суммированные площади А и Б соответствуют общей работе эластических компонентов, совершаемой при вдохе. Площадь Б соответствует всей работе неэластических компонентов, производимой при вдохе, то есть работе ткани и воздушной струи. Площадь Б соответствует работе, совершаемой неэластическими компонентами при выдохе Эта работа складывается из работы против эластических сил (см. рис. 17.26) и против неэластических (см. рис. 17.26). Для данного минутного объема существует интенсивность работы, при которой сумма эластического и зависимого от времени неэластического компонентов минимальна (рис. 17.27). При нормальном дыхании для перемещения воздуха в легкие и из них требуется менее 5% общего потребления кислорода (рис. 17.28). 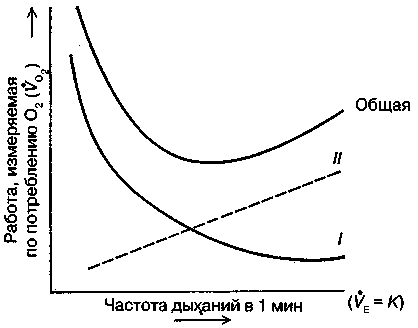 Рис. 17.27. Гипотетическая кривая работы эластических и неэластических компонентов и суммарной работы, производимой при равной частоте дыхания у человека при постоянном минутном объеме. Обратите внимание на оптимальную частоту, когда общая сумма работы эластических (I) и неэластических (II) компонентов дает минимум общей необходимой работы. Работа выражена в количестве потребляемого О2 (V02) 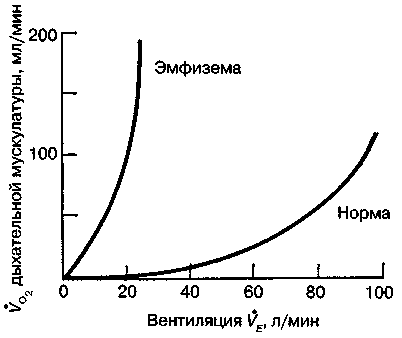 Рис. 17.28. Потребление кислорода дыхательными мышцами при разной частоте дыхания в норме и при пониженной растяжимости легких (эмфизема) Чем интенсивнее физическая работа, тем выше потребление кислорода дыхательной мускулатурой. J.M. Petit и др. (1962) установили зависимость между КПД и частотой дыхания у человека. Авторы регистрировали ЭМГ диафрагмы и прямой мышцы живота и сделали вывод, что при медленном и глубоком дыхании возникает дискоординация мышц-антагонистов, а при учащенном дыхании их функционирование было более согласованным. Именно этим фактором они объясняют увеличение КПД по мере учащения дыхания. А.В. Otis (1950) предложил определить механическую мощность дыхания при помощи следующего уравнения: W = K1V2+K2V3, где W — механическая мощность внешнего дыхания (Вт); V — минутный объем дыхания; К1и К2 — константы. Первая часть уравнения характеризует мощность, необходимую для преодоления эластического сопротивления легких и грудной клетки плюс ламинарного сопротивления воздушного потока в дыхательных путях; вторая часть — мощность, необходимую для преодоления турбулентного сопротивления потока воздуха в дыхательных путях. У человека в покое и при легкой физической работе с величинами МОД, не превышающими 30 л, механическая мощность внешнего дыхания составляет 0,04—0,31 Вт, однако при величине МОД 120—125 л эта мощность достигает 6,97—8,37 Вт. При увеличении МОД на 25 л по отношению к состоянию покоя (8—12л) кислородная стоимость дыхания увеличивается и на каждый литр вентиляции затрачивается дополнительно 1 мл кислорода (О2), а при возрастании МОД на 50—80 л — соответственно 2,0— 3,2 мл О2. Если величина МОД превышает 100 мл, на работу дыхательной мускулатуры затрачивается более 1 л О2. Если МОД превышает 150 л, то кислородная стоимость дыхания составляет около 4,5 л. R.J. Shepard (1966) считает, что уровень МОД в 120 л — это критическая граница, выше которой энергетическая стоимость работы аппарата внешнего дыхания становится особенно высокой. Изменения объема легких Объем легкого меняется при вдохе не всюду одинаково. Для этого имеются три главные причины. Во-первых, грудная полость во всех направлениях увеличивается неравномерно. Во-вторых, не все части легкого одинаково растяжимы. В-третьих, предполагается существование гравитационного эффекта, который способствует смещению легкого книзу (рис. 17.29). 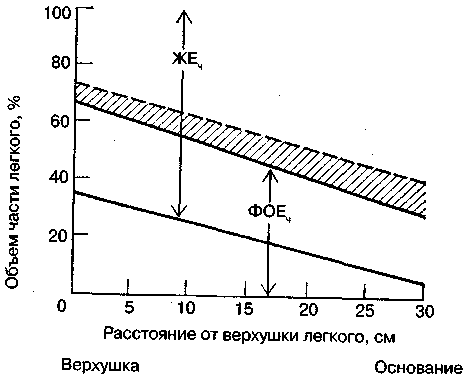 Рис. 17.29. Объем части легкого, способный к расширению, то есть жизненная емкость части (ЖЕч). Верхушка легкого показана в левой части графика ФОЕ — функциональная остаточная емкость Объем воздуха, вдыхаемый при обычном (неусиленном) вдохе и выдыхаемый при обычном (неусиленном) выдохе, называется дыхательным воздухом. Объем максимального выдоха после предшествовавшего максимального вдоха называется жизненной емкостью легких (ЖЕЛ). Она не равна всему объему воздуха в легком (общему объему легкого), поскольку легкие полностью не спадаются. Объем воздуха, который остается в неспавшихся легких, называется остаточным воздухом. Имеется дополнительный объем, который можно вдохнуть при максимальном усилии после нормального вдоха. А тот воздух, который выдыхается максимальным усилием после нормального выдоха, это резервный объем выдоха. Функциональная остаточная емкость состоит из резервного объема выдоха и остаточного объема. Это тот находящийся в легких воздух, в котором разбавляется нормальный дыхательный воздух. Вследствие этого состав газа в легких после одного дыхательного движения обычно резко не меняется. Минутный объем (V) — это воздух, вдыхаемый за одну минуту. Его можно вычислить, умножив средний дыхательный объем (V1) на число дыханий в минуту (f), или V = fVt. Часть Vt, например, воздух в трахее и бронхах до конечных бронхиол и в неперфузируемых альвеолах, не участвует в газообмене, так как не приходит в соприкосновение с активным легочным кровотоком — это так называемое мертвое пространство (Vd). Часть Vtкоторая участвует в газообмене с легочной кровью, называется альвеолярным объемом (Va). С физиологической точки зрения альвеолярная вентиляция (Уо) — наиболее существенная часть наружного дыхания Va = f(Vt— Vd), так как она является тем объемом вдыхаемого за минуту воздуха, который обменивается газами с кровью легочных капилляров. Вентиляция легких Вентиляция легких зависит от дыхательного объема (ДО) и частоты дыхания. Объем воздуха, который могут вместить легкие при максимально глубоком вдохе, называется общей емкостью легких (ОЕЛ). Тот объем, который человек может выдохнуть после максимального вдоха, составляет жизненную емкость легких (ЖЕЛ). Нормальная глубина дыхания, свойственная отдельному человеку в состоянии покоя, называется дыхательным объемом (ДО) и составляет около 10% ОЕЛ или 15—18% ЖЕЛ. Произведение дыхательного объема на число дыханий составляет минутный объем дыхания (МОД). Эта величина зависит прежде всего от уровня метаболизма, массы тела (веса), возраста, и в условиях покоя у взрослого человека может колебаться в широких пределах от 3 до 10 л. На рис. 17.30 схематично представлены легочные объемы человека. Вверху большая диаграмма показывает четыре первичных легочных объема и их примерную величину. Внешний круг указывает наибольший объем, до которого могут быть растянуты легкие; внутренний круг (остаточный объем) ограничивает объем, оставшийся после того, как весь воздух изгнан из легких (при самостоятельном дыхании). Вокруг центральной диаграммы расположены более мелкие; затушеванные области на них означают четыре емкости легких. Объем газа мертвого пространства включен в остаточный объем, функциональную остаточную емкость и общую емкость легких, как это имеет место при измерении обычными методами. Внизу представлены легочные объемы так, как они получаются на спирограмме; затушеванные участки соответствуют центральной диаграмме в верхней части рисунка. Из общего количества воздуха, вдыхаемого в нормальных условиях человеком, около 150 мл не попадает в альвеолы и распределяется в верхних дыхательных путях — глотке, гортани, трахее и бронхах — в так называемом мертвом пространстве (МП) и, следовательно, не участвует в газообмене. Различают анатомическое и физиологическое мертвое пространство. Объем анатомического мертвого пространства можно вычислить по формуле: ОМП (мл) = масса тела (кг) х 2,22. В обычных условиях величина анатомического МП довольно постоянна. 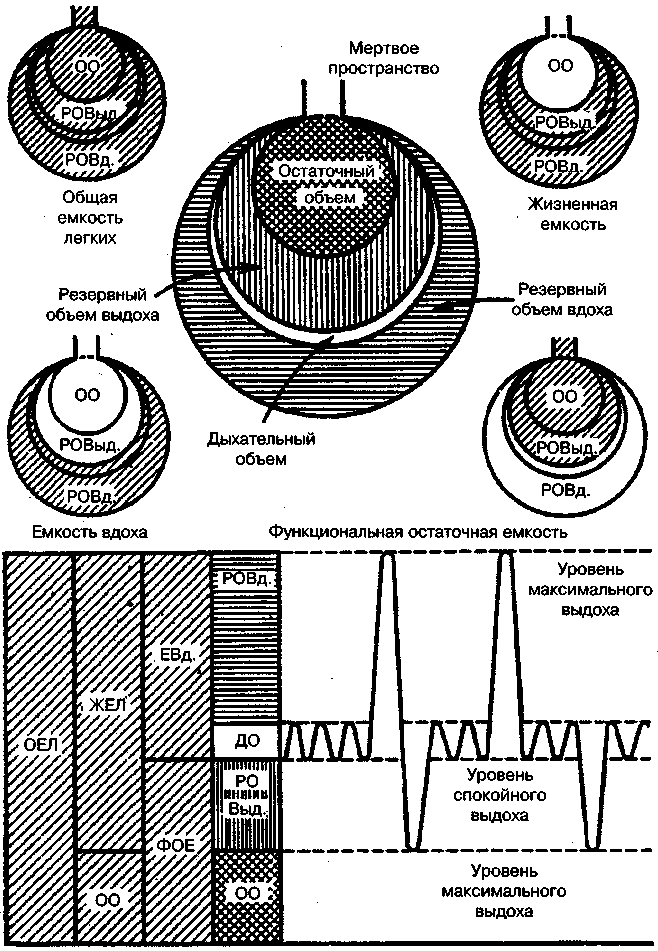 Рис. 17.30. Легочные объемы В процессе дыхания не весь вдыхаемый воздух достигает альвеол и участвует в газообмене; поэтому возникает необходимость введения еще одного понятия — минутной альвеолярной вентиляции (МАВ). У взрослого человека МАВ составляет в среднем 2,5— 5 л/мин. Зависимость между минутным объемом дыхания (МОД) и минутной альвеолярной вентиляцией может быть выражена формулами: МАВ = МОД — ОМП • ЧД или МАВ = (ДО — ОМП) • ЧД Поскольку МАВ определяет газообмен, уменьшение доли ее в МОД будет приводить к ухудшению газообмена и наоборот. При одном и том же МОД увеличение частоты дыхания (ЧД) приводит к снижению МАВ и, следовательно, к ухудшению газообмена. На рис. 17.31 показано, что один и тот же МОД (8000 мл) может быть получен при разной частоте дыхания (и, конечно, при разном ДО). Но если при нормальной ЧД и нормальном ДО доля альвеолярной вентиляции в МОД достаточно высока и составляет 5600 мл (см. рис. 17.31, б), то при тахипноэ МАВ снижается до 3200 мл, а доля объема, не участвующего в газообмене, увеличивается (см. 17.31, а). Это влечет за собой ухудшение газообмена и увеличение цены дыхания. Важным элементом адекватной спонтанной вентиляции легких здорового и больного организмов является синхронная деятельность межреберных дыхательных мышц и диафрагмы в активной фазе дыхательного цикла, т. е. в период вдоха, обеспечивающая максимальное увеличение емкости грудной полости в этот период. В ряде случаев наблюдается расстройство такой синхронизации в результате действия различных факторов. Такая синхронизированная деятельность дыхательных мышц и диафрагмы называется «наружным парадоксальным дыханием». Во всех случаях при парадоксальном дыхании возникают существенные нарушения газообмена, приводящие к гипоксии и гиперкапнии. На рис. 17.32 представлены возможные варианты такого нарушения вентиляции легких.  Рис. 17.31. Влияние изменений дыхательного объема и частоты дыхания на альвеолярную вентиляцию 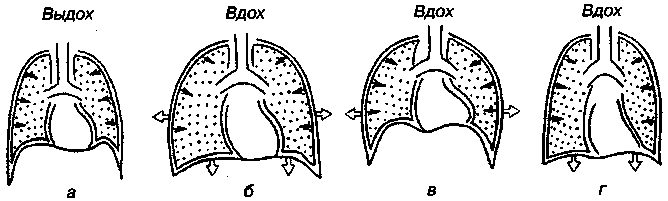 Рис. 17.32. Схема, иллюстрирующая «наружное парадоксальное дыхание»:а, б — взаимоотношение реберного каркаса грудной клетки и диафрагмы в момент нормального выдоха и вдоха; в — реберное дыхание; г — диафрагмальное дыхание Вентиляционно-перфузионные отношения. Легочный кровоток Эффективность газообмена в легких зависит от того, как распределяется объем вдыхаемого воздуха в альвеолах и кровоток в легочных сосудах. В идеальном случае на каждый метр протекающей по легочным сосудам крови в минуту должно приходиться 0,8 л альвеолярного воздуха, т. е. так называемый вентиляционно-перфузионный коэффициент равен 0,8 (рис. 17.33). Если проанализировать газообмен здорового человека, то почти во всех случаях будет обнаружена большая или меньшая неравномерность распределения воздуха в легких. У здорового человека в состоянии покоя в дыхании участвуют не все альвеолы, а в кровообращении — не все легочные капилляры. Однако равномерность распределения воздуха в легких возрастает с увеличением МОД, например, при физической нагрузке. 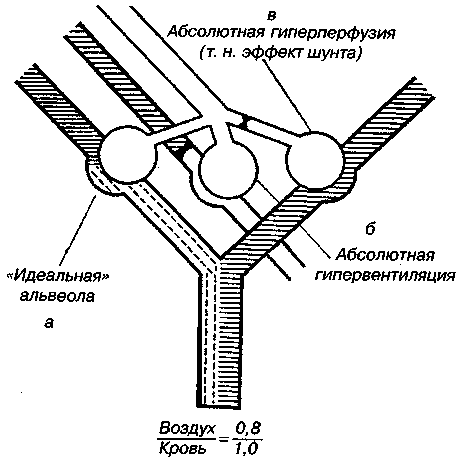 Рис. 17.33. Вентиляционно-перфузионные отношения в легких: а — нормальная вентиляция и нормальная перфузия; б — нормальная вентиляция альвеолы и отсутствие перфузии; в — отсутствие вентиляции и нормальная перфузия Неравномерное распределение кровотока по легочным сосудам также приводит к нарушению вентиляционно-перфузионных отношений. Даже у здорового человека почти никогда не бывает идеально равномерного распределения кровотока, так же как и вентиляция. При изменениях положения тела возникают изменения распределения кровотока в связи с гравитацией. Возникновение у неподвижных больных (особенно у больных в послеоперационном периоде при длительном пребывании в одном положении и др.) влажных, так называемых застойных хрипов в нижних задних отделах легких (при отсутствии их в верхних отделах) связано именно с неравномерным распределением кровотока и вентиляции. Тот факт, что насыщение артериальной крови О2 никогда не достигает 100% ив норме составляет 96%, объясняется неравномерностью вентиляции и кровотока, в результате которого в крови легочных вен всегда имеется небольшое количество восстановленного гемоглобина. Таким образом, в норме вентиляционно-перфузионные отношения каждого легкого в отдельности поддерживаются автономными механизмами в зависимости от ряда внешних и внутренних причин. Регуляция дыхания Известно, что главная функция легких состоит в обмене кислорода (О2) и углекислого газа (СO2) между воздухом и кровью, т. е. в поддержании нормальных уровней P О2и Рсо2 в артериальной крови. Уровни СО2 (Н+) и О2 в артериальной крови, как правило, регулируются в узких пределах через легочную вентиляцию. Несмотря на широкую изменчивость поглощения кислорода (О2) в организме и выделения из него углекислоты (СО2), Р0 и Рсов артериальной крови в норме сохраняются достаточно постоянными. Эта удивительная регуляция осуществляется благодаря тонкому управлению легочной вентиляцией. В ЦНС имеются специальные области, которые участвуют в создании каждого вентиляционного усилия дыхательных мышц, а также регулируют общую деятельность дыхательной системы. Участие ЦНС складывается из двух функционально раздельных элементов: 1) автоматическое дыхание, связанное, главным образом, со структурами ствола мозга, и 2) произвольное дыхание, связанное со структурами высших уровней мозга, главным образом, с корой больших полушарий. Выяснено, что высший отдел ЦНС, кора больших полушарий, оказывает влияние на глубину и частоту дыхания. При стимуляции особых областей коры головного мозга дыхание или усиливается, или ослабляется. Эти области находятся под произвольным контролем и проявляют себя, когда мы едим или говорим. Система регуляции дыхания (рис. 17.34) включает три основных элемента: 1) рецепторы, воспринимающие информацию и передающие ее в 2) центральный регулятор, расположенный в головном мозге. Здесь информация обрабатывается и отсюда же посылаются команды на: 3) эффекторы (дыхательные мышцы), непосредственно осуществляющие вентиляцию легких.  Рис. 17.34. Основные элементы системы регуляции дыхания. Информация от различных рецепторов поступает в центральный регулятор, а от него посылаются команды дыхательным мышцам. Изменение активности этих мышц приводит к изменению вентиляции, а это в свою очередь снижает возбуждающие воздействия на рецепторы (отрицательная обратная связь) Кроме того, существует ряд механорецепторов, возбуждение которых влияет на характер дыхания. Среди них — рецепторы давления. При их возбуждении возникают реакции, варьирующие от временного апноэ до значительного учащения дыхания. Движение суставов и растягивание мышц конечностей повышают как частоту дыхания, так и дыхательный объем. Боль тоже действует на дыхание. Реакция легких на физические нагрузки Пока внутриплевральное давление остается ниже атмосферного, размеры легких точно следуют за размерами грудной полости. Движения легких совершаются в результате сокращения дыхательных мышц в сочетании с движением частей грудной стенки и диафрагмы. Вентиляция и легочный кровоток, перенос О2 и СО2 и диффузная способность при физической нагрузке могут возрастать в несколько раз. При физической нагрузке вентиляция легких резко возрастает и при интенсивной физической работе может становиться очень сильной. У здоровых молодых мужчин максимальное потребление кислорода (МПК) иногда достигает 4 л/мин, а легочная вентиляция — 120 л/мин, т. е. в 15 раз превышает уровень покоя. Усиление вентиляции тесно связано с увеличением потребления О2 и выделения СО2. Интересно, что причины такого усиления при физической нагрузке еще во многом неясны (J.B. West, 1988). При нагрузке Рсо2в артериальной крови не увеличивается; напротив, при тяжелой физической работе оно обычно слегка снижается. При умеренной нагрузке рН артериальной крови остается почти постоянным, а при тяжелой физической работе — снижается в связи с выделением молочной кислоты (лактата) в процесс анаэробного гликолиза. Ясно, что ни один из перечисленных факторов не должен вызывать резкого усиления вентиляции при легкой или умеренной физической нагрузке. Исследования показывают, что если совершать пассивные движения конечностями, то вентиляция легких усиливается. Это, по-видимому, связано с рефлекторной реакцией рецепторов, расположенных в суставах или мышцах. Предполагается, что увеличение вентиляции легких при физической нагрузке может быть частично обусловлено повышением температуры тела и импульсами, поступающими от двигательной коры головного мозга. Исследования показывают, что тренировки (и, особенно, соревнования) в среднегорье и зонах жаркого и влажного климата, вызывают сильную реакцию организма спортсмена на внешние факторы. |