ответы на физиологию человека. 24-26 Физиология. Вопросы к контрольной
 Скачать 83.58 Kb. Скачать 83.58 Kb.
|

|
Вопросы к контрольной. 1. Возбудимые ткани. Характеристика возбудимых тканей. Общие свойства (возбудимость, раздражимость, проводимость, лабильность). 2. Возбудимость как высокодифференцированная специализированная форма раздражимости. Способы оценки возбудимости. Мера возбудимости – порог раздражения. 3. Электрические явления в возбудимых тканях. История их открытия (Л. Гальвани, Э. Дюбуа-Реймон). 4. Современные представления о процессе возбуждения. Возбуждение как биологическая реакция. Мембранно-ионная теория возбуждения. 5. Методы изучения и современные представления о строении и функции клеточной мембраны. 6. Мембранный потенциал. Роль концентрационных градиентов и избирательной проницаемости мембраны в возникновении мембранного потенциала. 7. Потенциал действия и его фазы. Критический уровень деполяризации. Местный процесс (локальный ответ) и переход его в распространяющийся потенциал – потенциал действия. 8. Динамика возбудимости в процессе формирования потенциала действия. 9. Законы раздражения возбудимых тканей. 10. Понятие о реобазе и хронаксии. Аккомодация возбудимых тканей. 11. Физиологические свойства мышечной ткани. 12. Скелетные мышечные волокна. Морфологические особенности «быстрых» и «медленных» мышечных волокон. 13. Сократимость как специфическое свойство мышечной ткани. Сокращение мышцы, методика регистрации. Электромиография. Клиническое значение электромиографии. Изометрический и изотонический режим сокращения мышц. Электронно-микроскопическое строение мышцы. 14. Механизм мышечного сокращения. Роль потенциала действия, ионов кальция и АТФ в механизме мышечного сокращения. 15. Явления, сопровождающие мышечное сокращение (химические, тепловые, биоэлектрические). 16. Одиночное сокращение мышцы и его фазы. Изменения возбудимости мышцы в процессе одиночного сокращения. Влияние силы раздражения на величину сокращения мышцы. 17. Суммация сокращений и тетанус. Механизм тетануса (Л. Гельмгольц, Н. Е. Введенский). 18. Оптимум и пессимум частоты раздражения. Лабильность. 21.Физиологические свойства гладких мышц (возбудимость, проводимость, сократимость, пластичность, автоматия). Сравнение свойств гладких и поперечно-полосатых мышц. Особенности нервно-мышечной передачи возбуждения в гладких мышцах. 22. Физиологические свойства нервных волокон. (возбудимость, проводимость). 23. Структурно-функциональная характеристика нервных волокон (миелиновых и безмиелиновых). 24. Классификация нервных волокон. Волокна типа A, B, C. Скорость проведения возбуждения в различных нервных волокон. 25. Законы проведения возбуждения в нервных волокон. 26. Механизм проведения возбуждения в мякотных (миелиновых) и безмякотных (безмиелиновых) нервных волокнах – сальтаторный и непрерывный. 27. Параметры функционального состояния нерва (порог раздражения, лабильность, полезное время, реобаза, хронаксия). Закон Дюбуа Реймона. 28. Ультраструктура нервно-мышечного синапса. 29. Функциональные свойства и особенности нервно-мышечного синапса. Одностороннее проведение возбуждения в синапсах. Синаптическиая задержка. Быстрая «утомляемость» синапсов 24. Классификация нервных волокон. Волокна типа A, B, C. Скорость проведения возбуждения в различных нервных волокон. А – нервные волокна с самой толстой миелиновой оболочкой. Наиболее высокая скорость передачи нервного импульса. В – миелиновая оболочка тоньше, скорость проведения возбуждения ниже +С – безмиелиновые волокна с относительно низкой скоростью передачи импульса. 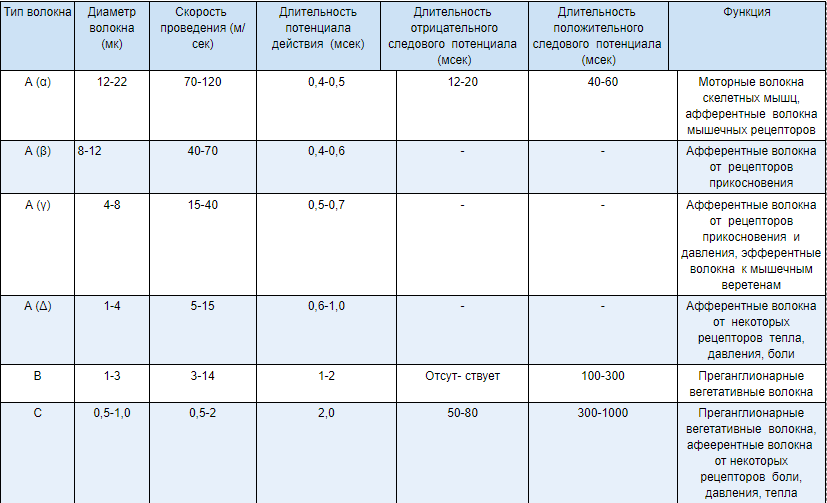 Проведение возбуждения по немиелинизированным волокнам В безмиелиновых волокнах нервный импульс распространяется волнообразно, последовательно возбуждаются небольшие участки размером в несколько микрометров. Возбуждение распространяется гораздо медленнее, чем по миелиновым волокнам. Внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, так как липидная мембрана имеет высокое электрическое сопротивление. Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда. На границе возбужденного и невозбужденного участка начинает протекать электрический ток. Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения, в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 2, Г). Таким образом, волна возбуждения охватывает все новые участки мембраны нервного волокна. При действии раздражителя пороговой силы на мембрану безмиелинового волокна изменяется ее проницаемость для ионов Nа+, которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны (внутренняя становиться заряженной положительно, а наружная отрицательно). Это ведет к возникновению круговых токов (заряженных частиц) от «+» к «–» на протяжении всего волокна. Особенности распространения возбуждения по безмиелиновым волокнам: 1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением. 2. Возбуждение распространяется с небольшой скоростью. 3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна). По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров. Проведение возбуждения по миелинизированным волокнам Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. В миелинизированном волокне токи проводятся только в зонах, не покрытых миелином (перехватах Ранвье). Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением. При действии раздражителя пороговой силы на мембрану миелинового волокна в области перехвата Ранвье изменяется проницаемость для ионов Nа+, которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны, что ведет к возникновению круговых токов. Этот ток идет через межтканевую жидкость к соседнему перехвату, где происходит смена заряда. Таким образом, возбуждение перепрыгивает с одного участка на другой. Обратное движение возбуждения невозможно так как участок, через который оно прошло, находится в фазе абсолютной рефрактерности. Особенности распространения возбуждения по миелиновым волокнам: 1. Распространение ПД в миэлинизированных нервных волокнах осуществляется сальтаторно - скачкообразно от перехвата к перехвату, т.е. возбуждение (ПД) как бы «перепрыгивает» через участки нервного волокна, покрытые миелином, от одного перехвата к другому и все волокно сразу не охватывается возбуждением. 2. Возбуждение распространяется с большой скоростью. 3. Возбуждение распространяется без декримента. По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, т.е. там, где требуется высокая скорость ответной реакции. В миелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только в участках мембраны, расположенных в области перехватов Ранвье. +При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис. 3, А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис. 3, Б). Однако в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис. 3, В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому. 25. Законы проведения возбуждения по нервам. Нервное волокно обладает следующими физиологическими свойствами: возбудимостью, проводимостью, лабильностью. Проведение возбуждения по нервным волокнам осуществляется по определенным законам. Закон двустороннего проведения возбуждения по нервному волокну. Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от возбужденного участка (места его возникновения), т. е., центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения. Естественным направлением распространения возбуждения является: в афферентных проводниках - от рецептора к клетке, в эфферентных - от клетки к рабочему органу. Закон анатомической и физиологической целостности нервного волокна. Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву (законы анатомической и физиологической целостности). Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е., к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается. Закон изолированного проведения возбуждения по нервному волокну. В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервного волокна на другое, то нормальное функционирование органов было бы невозможно. Возбуждение (потенциал действия) распространяется по нервному волокну без затухания. Периферический нерв практически неутомляем. 26. Механизм проведения возбуждения по нерву. Возбуждение (потенциал действия - ПД) распространяется в аксонах, телах нервных клеток, а также иногда в дендритах без снижения амплитуды и без снижения скорости (бездекрементно). Механизм распространения возбуждения у различных нервных волокон неодинаков. При распространении возбуждения по безмиелиновому нервному волокну механизм проведения включает два компонента: раздражающее действие катэлектротона, порождаемое локальным ПД, на соседний участок электровозбудимой мембраны и возникновение ПД в этом участке мембраны. Локальная деполяризация мембраны нарушает электрическую стабильность мембраны, различная величина поляризации мембраны в смежных ее участках порождает электродвижущую силу и местный электрический ток, силовые линии которого замыкаются через ионные каналы. Активация ионного канала повышает натриевую проводимость, после электротонического достижения критического уровня деполяризации (КУД) в новом участке мембраны генерируется ПД. В свою очередь этот потенциал действия вызывает местные токи, а они в новом участке мембраны генерируют потенциал действия. На всем протяжении нервного волокна происходит процесс новой генерации потенциала действия мембраны волокна. Данный тип передачи возбуждения называется непрерывным. Скорость распространения возбуждения пропорциональна толщине волокна и обратно пропорциональна сопротивлению среды. Проведение возбуждения зависит от соотношения амплитуды ПД и величины порогового потенциала. Этот показатель называется гарантийный фактор(ГФ) и равен 5 - 7, т.е. ПД должен быть выше порогового потенциала в 5- 7 раз. Если ГФ = 1 проведение ненадёжно, если ГФ < 1 проведения нет. Протяженность возбуждённого участка нерва L является произведение времени (длительности) ПД и скорости распространения ПД. Например, в гигантском аксоне кальмара L= 1 мс25 мм/мс = 25 мм. Наличие у миелиновых волоконоболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. Вмиелинизированном волокне токи проводятся только в зонах, не покрытых миелином, - перехватах Ранвье, в этих участках и генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением. Распространение ПД в миэлинизированных нервных волокнах осуществляетсясальтаторно- скачкообразно от перехвата к перехвату, т.е. возбуждение (ПД) как бы «перепрыгивает» через участки нервного волокна, покрытые миелином, от одного перехвата к другому. Скорость такого способа проведения возбуждения значительно выше, и он более экономичен по сравнению с непрерывным проведением возбуждения, поскольку в состояние активности вовлекается не вся мембрана, а только ее небольшие участки в области перехватов, благодаря чему уменьшается нагрузка на ионный насос. 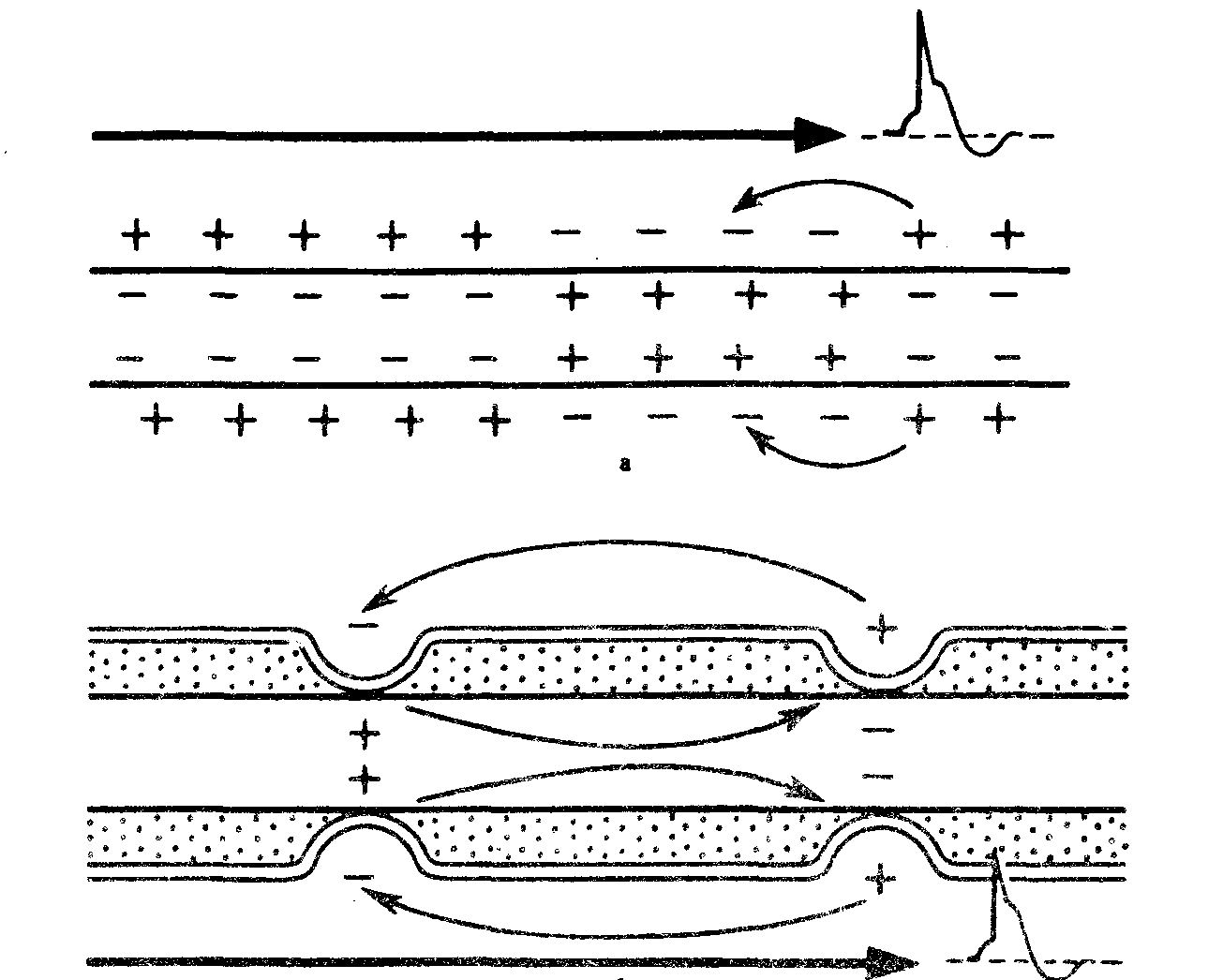 +Схема распространения возбуждения в безмиелиновых и миелиновых нервных волокнах. |