Коллоквиум по возбудимым тканям. Вопросы коллоквиума по разделу Физиология возбудимых тканей
 Скачать 0.85 Mb. Скачать 0.85 Mb.
|

|
4) Удаление АХ из синапт. щели. Оно осущ. путём разрушение под действием его ингибитора – АХЭ. Около 60% холина захватываектся обратно пресинапт. окончанием, что делает синтез медиатора более экономичным, часть АХ рассеивается. В промежутках между ПД из пресинапт. окончания спонтанно выделяются 1-2 кванта медиатора в синапт. щель в течение секунды, формируя так наз. миниатюрные потенциалы = 0,4-0,8 мВ. Они поддерживают высокую возбудимость иннервируемой клетки в условиях функц. покоя и играют троф. роль, а в ЦНС они способствуют поддержанию тонуса её центров. Если распад медиатора или обратный захват замедляется, то действие выделившегося медиатора усиливается и становится более продолжительным. Это имеет клиническое значение. Например, действие многих антидепрессантов и антипаркинсонических средств заключается в торможении обратного захвата медиаторов. 18. Механизмы и пути блокирования передачи возбуждения в мионевральном синапсе. Пути блокады. При блокаде скелетная мышцы расслабляется – это осущ. миорелаксанты. Блокируют передачу возбуждения: 1) Вещ-ва блокаторы холинрецптора 2) Нарушение синтеза АХ 3) Вытеснение АХ из рецептора 4) Инактивация постсинаптической мембраны 4) АХЭ-ные вещ-ва. 19. Понятие о миорелаксантах, их применение в медицинской практике. М  иорелаксанты — хим. соединения, обладающие N-холинолитическими свойства и избирательно блокирующими передачу в неврно-мыш. синапсе, что ведёт к расслаблению поперечно-полосатой мускулатуры. иорелаксанты — хим. соединения, обладающие N-холинолитическими свойства и избирательно блокирующими передачу в неврно-мыш. синапсе, что ведёт к расслаблению поперечно-полосатой мускулатуры.Анализируя механизм воздействия на организм человека, можно различить всего два вида: препараты периферического действия и центральные миорелаксанты. Общие показания к применению миорелаксантов. 1. Обеспечение условий для интубации трахеи. 2. Обеспечение миорелаксации во время оперативных вмешательств для создания оптимальных условий работы хирургической бригады без избыточных доз препаратов для общей анестезии, а также необходимость мышечного расслабления при некоторых диагностических манипуляциях, выполняемых в условиях общей анестезии (например, бронхоскопия). 3. Подавление самостоятельного дыхания с целью проведения ИВЛ. 4. Устранение судорожного синдрома при неэффективности противосудорожных препаратов. 5. Блокада защитных реакций на холод в виде мышечной дрожи и гипертонуса мышц при искусственной гипотермии. 6. Миорелаксация при репозиции отломков костей и вправлении вывихов в суставах, где имеются мощные мышечные массивы. Общие противопоказания к применению миорелаксантов. 1. Миастения, миастенический синдром, миопатия Дюшена или Беккера, миотония, семейные периодические параличи. 2. Отсутствие условий для проведения ИВЛ. Побочные эффекты после приема. Снижение концентрации внимания, что наиболее опасно для людей, сидящих за рулем автомобиля. Понижение артериального давления. Повышенная нервная возбудимость. Ночное недержание мочи. Аллергические проявления. Проблемы со стороны ЖКТ. Судорожные состояния. 20. Физиологические особенности свойств скелетных мышц. Скелетная (поперечнополосатая) мышечная ткань — упругая, эластичная ткань, способная сокращаться под влиянием нервных импульсов: один из типов мышечной ткани. П-п. мышцы не способны работать без управляющей импульсации из ЦНС. Физиологические свойства скелетных мышц: 1)Возбудимость –способность отвечать на раздражения возникновением ПД. Ниже, чем возбудимость нервной ткани, что объясняется низкой величиной мембранного потенциала. Возбуждение распространяется вдоль мышечного волокна. 2) Проводимость – способность к проведению ПД вдоль мышечного волокна. Меньше проводимости нервной ткани (скелетная = 1-13 м/с). 3) Рефрактерный период - период времени после возникновения на возбудимой мембране потенциала действия, в ходе которого возбудимость мембраны снижается, а затем постепенно восстанавливается до исходного уровня. Рефрактерность ышечной ткани более продолжителен, чем нервной ткани. Длительность рефрактерного периода нерва 14 м/с, а поперечно-полосатой мышцы около 35 м/с. 4) Лабильность мышечной ткани значительно ниже, чем нервной. Действительно, икроножная мышца лягушки может воспроизводить 200 -250 волн возбуждения в 1 с, а седалищный нерв -500 -1000. 5) Сократимость – способность мышечного волокна изменять свою длину и степень напряжения в ответ на раздражение пороговой силы. Даже при спокойной ходьбе трехглавая мышца голени может развивать напряжение, которое почти в 4 раза больше веса идущего человека, а при беге икроножная мышца может развить напряжение, в 6 раз превышающее вес тела. Если бы все мышцы, содержащие примерно около 300 млн. волокон, возбуждались одновременно и максимально и осуществляли тягу в одном направлении, они смогли бы развить силу, по крайней мере, в 25 тонн. 6) Сокращение возможно благодаря растяжимости мышцы – способность изменять свою длину под действием растягивающей ее силы, и эластичности - способности мышцы принимать свою первоначальную длину после прекращения действия растягивающей или деформирующей силы. 7) Вязкость. Скелетные мышцы составляют 40% от массы тела и выполняют ряд важных функций: 1 - передвижение тела в пространстве, 2 - перемещение частей тела относительно друг друга, 3 - поддержание позы, 4 - передвижение крови и лимфы, 5 - выработка тепла, 6 - участие в акте вдоха и выдоха, 7 - двигательная активность как важнейший антиэнтропийный и антистрессовый фактор (тезисы " движение - это жизнь" или " кто много двигается, тот много живет" - имеют реальную материальную основу), 8 - депонирование воды и солен, 9 - защита внутренних органов (например, органов брюшной полости). 21. Особенности строения мембраны и саркомеров волокон скелетной мышцы. Механизм мышечного сокращения. С 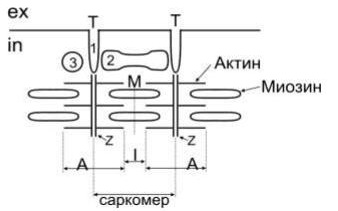 труктурной и сократит. единицей мышцы явл. мыш. волокно – сильно вытянутое многоядерное волокно. Толщина 10-100 мкм. Длина от ..мм до …см.М труктурной и сократит. единицей мышцы явл. мыш. волокно – сильно вытянутое многоядерное волокно. Толщина 10-100 мкм. Длина от ..мм до …см.М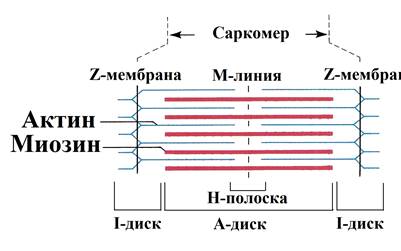 ыш. волокно имеет мембранную оболочку – сарколемму. В саркоплазме есть ядра, митохондрии, растворимые белки, капельки жира, гранулы гликогена, фосфатсодержащие вещ-ва и др. Там же расположены многочисленные мембраны. Они вкл. в состав Т-трубочек, пересекающих мыш. волокно и связывающих сарколемму с внутриклет. пространством и саркоплазматич. ретикулумом. Через Т-трубочки может также происходить выделение продуктов обмена из мыш. клетки в межклет. пространство и далее в кровь. ыш. волокно имеет мембранную оболочку – сарколемму. В саркоплазме есть ядра, митохондрии, растворимые белки, капельки жира, гранулы гликогена, фосфатсодержащие вещ-ва и др. Там же расположены многочисленные мембраны. Они вкл. в состав Т-трубочек, пересекающих мыш. волокно и связывающих сарколемму с внутриклет. пространством и саркоплазматич. ретикулумом. Через Т-трубочки может также происходить выделение продуктов обмена из мыш. клетки в межклет. пространство и далее в кровь.Внутри мыш. волокна от одного конца до другого тянутся миофибриллы (d=1-2 мкм), 1 мыш. волокно =2000 миофибрилл. Миофибриллы группируются в пучки по 4-20 шт. В одиночной миофибрилле насчитывается 2000-25000 протофибрилл – параллельно лежащих нитей 2х типов – толстых и тонкий. Толстые нити суст. из миозина, тонкие – из актина. В продольных борозках актиновых нитей мофибрилл расположены так наз. регуляторные единицы, состоящие из белков тропомиозина и торпонина. Эти белки в невозбуждённой мышце блокируют взаимодействие между актином и миозином. Миофибриллам свойственна поперечная исчерченность – чередование тёмных и светлых полос. Тёмные полосы в поляризованном свете проявляют св-ва двойного лучепреломления и поэтому наз. А-дисками (анизотропными). Это место расположения группы параллельных толстых миофибрилл, которая чередуется с изотропной группой тонких протофибрилл – I-диски. Светлые I-диски делятся поплам поперечной тёмной Z-линией. Участок между 2мя линиями – саркомер (l=2-3 мкм). В каждом саркомере есть 2 набора тонких нитей, прикреплённых к Z-линии, и один комплекс толстых нитей, сосредоточенных в А-диске. Миозиновые нити имеют выступы – поперечные мостики. Они отходят от нитей биполярно. Поперечные мостики, состоящие из головки и шейки, в спокойном состоянии не могут соединиться с актиновыми нитями из-за особого расположения тропомиозина, закрывающего активные центры актина и препятствующего их взаимодействию с поперечными мостиками миозина. Тропонин подавляет миозин-АТФ-азную активность, что делает невозможным расщепление АТФ, т.е. мыш. волокна пребывают в расслабленном состоянии. Сокращение мыш. волокон первично связано с процессами генерации и распр. ПД по поверхностной мембране, а также по мембранам, выстилающим поперечные трубочки Т-системы. Проникая внутрь волокна, ПД за счёт своего эл. поля приводит к деполяризации мембран продольных трубочек и цистерн саркоплазматического ретикулума. Снижение их ПП вызывает выход Са2+ из боковых цистерн в межфибриллярное пространство. Свободный Са2+ запускает процесс взаимодействия актина с миозином и сокращения мышцы. Совокупность явлений, обуславливающих связь между возбуждением и сокращением мыш. волокон – электромех. сопряжение. М 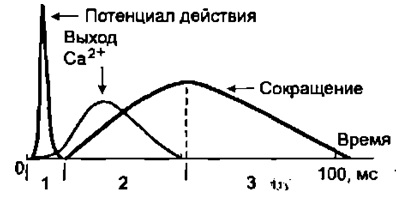 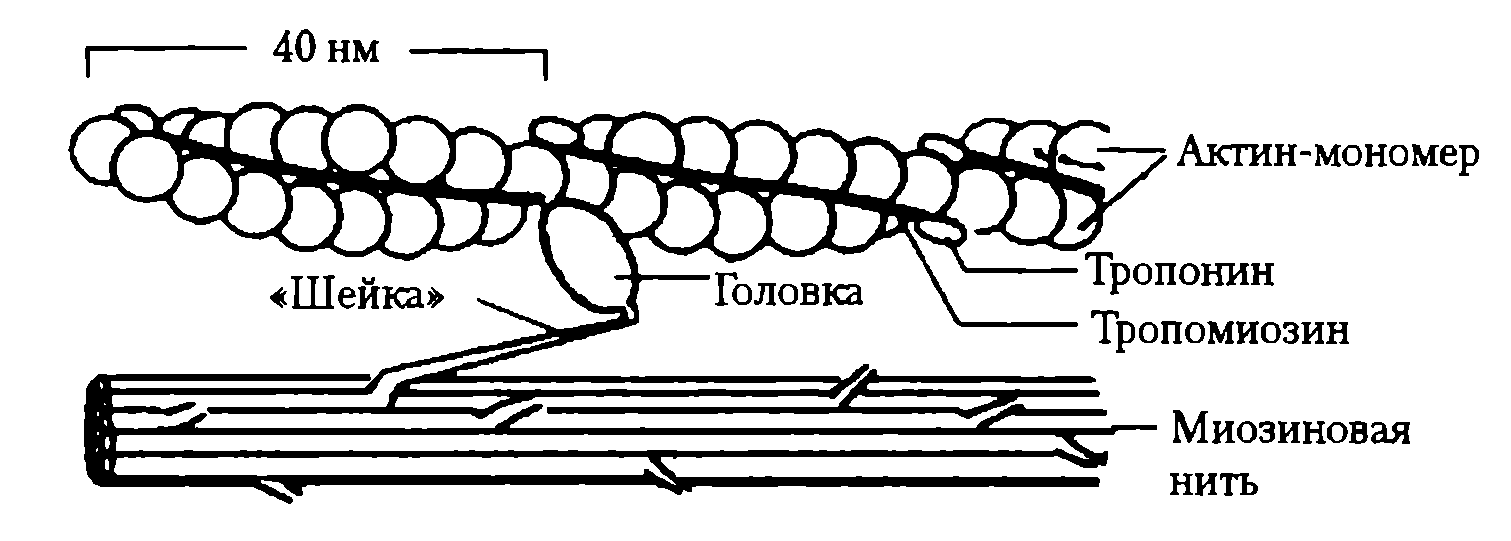 еханизм. В присутствии ионов Са2+, а также АТФ пропонин изменяет свою конфигурацию и отодвигает нить тропомиозина, обеспечивая возможность соединения головки поперечного мостика миозина с актином. Соединение головки фосфорилированного миозина с актином приводит к тому, что головка приобретает АТФазную активность, в ней происходит гидролиз АТФ, сопровождаемый изменением пространственной ориентации. Последнее носит форму гребкового движения, обеспечивающего втягивание тонких актиновых миофиламентов в промежутки между толстыми миозиновыми миофиламентами на один шаг без изменения длины миофиламента. Затем следует отрыв мостика и повторение всего цикла. При каждом гребковом движении головки поперечного мостика расщепляется одна молекула АТФ. еханизм. В присутствии ионов Са2+, а также АТФ пропонин изменяет свою конфигурацию и отодвигает нить тропомиозина, обеспечивая возможность соединения головки поперечного мостика миозина с актином. Соединение головки фосфорилированного миозина с актином приводит к тому, что головка приобретает АТФазную активность, в ней происходит гидролиз АТФ, сопровождаемый изменением пространственной ориентации. Последнее носит форму гребкового движения, обеспечивающего втягивание тонких актиновых миофиламентов в промежутки между толстыми миозиновыми миофиламентами на один шаг без изменения длины миофиламента. Затем следует отрыв мостика и повторение всего цикла. При каждом гребковом движении головки поперечного мостика расщепляется одна молекула АТФ.Последовательность событий в цикле сокращение-расслабление мыш. волокна: 1) поступление ПД по нервному волокну к мионевр. синапсу. 2) синаптич. активация мыш. волокна (возникновение ПКП и ПД). 3) проведение ПД вдоль клеточной мембраны и вглубь волокна по Т-трубочкам. 4) высвобождение Са2+ из бок. цистерн саркоплазм. ретикулума и диффузия его к миофибриллам. 5) конформация тропонин-тропомиозин. комплекса и контакт поперечных мостиков миозина с актином. 6) высвобождение энергии АТФ и скольжение актиновых и миозиновых нитей, выражающееся в укорочении миофибриллы. Далее активируется кальциевый насос, снижается конц. свободных ионов Са2+ в саркоплазме, в результате расслабляются миофибриллы. Энергии АТФ исп.: 1) на контакт актина и миозина, скольжения актиновых и миозиновых нитей, приводящий к укорочению миофибрилл. 2) на отрыв головки. 3) на работу Са-насоса, активируемого при расслаблении волокна. 4) на работу Na/К-насоса, обеспечивающего поддержание постоянства градиента конц. этих ионов. 22. Понятие двигательной единицы, физиологические особенности быстрых и медленных двигательных единиц. Комплекс мотонейрона со всеми его коллатералями и группой иннервируемых им мыш. волокон наз. двигательной единицей (ДЕ). Все мыш. волокна, относящиеся к одной ДЕ, рассеяны по всей мышце и независимо от их числа относят к одному типу и иннервируются только одной ветвью одного мотонейрона. Функциональное состояние ДЕ оценивают методом электромиографии (ЭМГ) – регистрации эл. активности интактной мышцы с помощью введённых в мышцу или приложенных к ней электродов. В абсолютной расслабленной мышце эл. активность почти отсутствует. При небольшом напряжении ДЕ раздражаются с невысокой частотой (5-10 имп/с), при большом – 20-30 имп/с. Обычным режимом естественной сократит. активности мыш. волокон ДЕ явл. зубчатый тетанус или даже ряд последовательных сокращений. Однако форма сокращения целой мышцы в этой ситуации напоминает гладкий тетанус. Причина этого – асинхронность разрядов мотонейронов и асинхронность сократительной реакции отдельных мыш. волокон, вовлечение большого х кол-ва, вследствие чего мышца плавно сокращается и плавно расслабляется. Функционально ДЕ разделяются на медленные и быстрые. 1) Медленные мотонейроны низкопороговые; к ним относятся в основном малые мотонейроны. По мере увеличения силы сокращения мышцы частота импульсации этих волокон изменяется незначительно. Медленные мотонейроны способны поддерживать длительный разряд без заметного снижения частоты импульсации на протяжении длит. отрезков времени, поэтому их наз. малоутомляемыми. 2) Быстрые мотонейроны высокопороговые. Они вкл. только для обеспечения относительно больших по силе стат. и динам. сокращений мышц, а также в начале любых сокращений, чтобы увеличить скорость нарастания напряжения мышцы или сообщить движущейся части тела необходимое ускорение. Чем больше скорость и сила движений, тем больше кол-во быстрых ДЕ в них участвуют. Утомляемы, т.к. не способны к длит. поддержке высокочастотного разряда. Быстрые мыш. волокна белые, более толстые, содержат больше миофибрилл и обладают большей силой, окружены меньшим кол-вом капилляров, в них меньше митохондрий, миоглобина жиров. Активность окислит. ферментов ниже, чем в медленных, однако активность гликолитических ферментов, запасы гликогена выше. Эти волокна не обладают большой выносливостью и более приспособлены для мощных, но относительно кратковременных сокращений. Красные волокна окружены богатой капиллярной сетью, позволяющей им получать большое кол-во О2 в крови. Повышенное содержание миоглобина облегчает транспорт О2 в мыш. клетках к митохондриям. Волокна содержат большое кол-во митохондрий и жиров, т.о. использование медленными мыш. волокнами более эффективного аэробного окислит. пути энергопродукции, т.о. высокая выносливость. Также есть тонические мыш. волокна, на них локализуются по 7-10 синапсов, принадлежащих, как правило, нескольким мотонейронам. ПКП этих мыш. волокон не вызывают в них ПД, а непосредственно запускают мыш. сокращение. 23. Энергетика мышечного сокращения. Пути ресинтеза АТФ. Мощность и емкость энергетических систем организма. Для активной деятельности мышцы необходим постоянный ресинтез АТФ. Запас АТФ в скелетных мышцах невелик — всего примерно на 10 одиночных сокращений. 1. Ферментативный перенос фосфатной группы от богатого энергией креатинфосфата на аденозиндифосфорную кислоту — эффективный путь, позволяющий за несколько секунд совершить большую работу, которая выполняется, например, спринтером или штангистом. В случае интенсивной мышечной работы запасы креатинфосфата быстро истощаются и реализуются другие, более медленные способы пополнения АТФ. 2. Гликолитический путь, связанный с анаэробным расщеплением глюкозы до молочной кислоты, позволяет синтезировать 2 молекулы АТФ на 1 молекулу глюкозы. Этот способ ресинтеза АТФ идет быстро, но накапливающаяся молочная кислота тормозит активность гликолитических ферментов. Такой вид энерголродукции используется при больших, но непродолжительных нагрузках, например у спортсменов при беге на средние дистанции, и способствует сдвигам кровообращения в работающей мышце для обеспечения адекватного хода третьего типа ресинтеза АТФ. 3. Аэробное окисление глюкозы, жирных кислот и аминокислот в цикле Кребса, совершаемое в митохондриях, — процесс экономичный: из одной молекулы глюкозы образуется около 38 молекул АТФ, а при окислении одной молекулы жирной кислоты — коло 128 молекул АТФ. Для получения энергии таким способом требуется больше времени, чем при первых двух способах, поэтому третий путь ресинтеза АТФ используется во всех случаях, когда мощность хватательной активности мышцы невысокая. Аэробное окисление глюкозы и жирных кислот — наиболее типичныйспособ энергообеспечения. Однако при длительной работе в организме накапливаются недоокисленные продукты и создаётся кислородная задолженность. Такой долг погашается после работы за счёт компенсаторной мобилизации кровообращения и дыхания (тахикардия, одышка). Если же работы, несмотря на наличие кислородного долга, продолжается, наступает выраженное состояние утомления, которое иногда прекращается за счёт мобилизации дополнительных резервов кровообращения и дыхания. 24. Характеристика видов и режимов мышечного сокращения: одиночное и тетаническое сокращение. Механизм тетанического сокращения. 2 основных типа сокращения. Сокращение мышцы, при котором её волокна укорачиваются, но напряжение остаётся постоянным, наз. изотоническим. При изомерическом сокращении мышцы укоротиться не может, если оба её конца закреплены неподвижнео. В этом случае по мере развития сократительного процесса напряжение возрастает, а длина мышечных волокон остаётся неизменной. В естественных условиях смешанное сокращение. Одиночное сокращение. В эксперименте возникает при раздражении мышцы одиночным эл. стимулом. Волна возбуждения возникает в месте приложения электродов для прямого раздражения мышцы или в области нервно-мыш. соединения и отсюда распр. вдоль всей мышцы. В естественных условиях сокращение мыш. волокна происходит при поступлении к нему нерв. импульса в области нерв.-мыш. синапса. Одиночной сокращение икроножной мышцы лягушки в изотон. режиме начинается через короткий скрытый (латентный) период – до 0,01 с, далее следует фаза укорочения – 0,05 с и фаза расслабления – 0,05-0,06 с. Обычно мышца укорач. на 5-10%. Продолжительность ПД мыш. волокон – до 10 мс, с учётом замедления реполяризации. Одиночное мыш. волокно реагирует на раздражение по правилу «всё или ничего», т.е. отвечает на все надпороговые раздражения стандартным ПД и стандартным одиночным сокращением. При увеличении силы раздражения число возбуждённых волокон растёт, пока все волокна е окажутся сокращёнными, и тогда достигается макс. сокращение мышцы. В 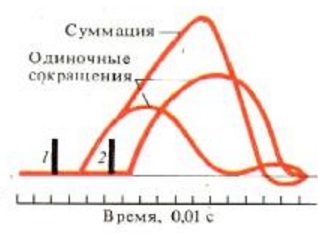 естественных условиях мыш. работают в режиме одиночных сокращений только при относит. низкой частоте импульсации мотонейров, когда интервалы между последовательными ПД превышают длительность одиночного сокращения, иннервируемых ими мыш. волокон. естественных условиях мыш. работают в режиме одиночных сокращений только при относит. низкой частоте импульсации мотонейров, когда интервалы между последовательными ПД превышают длительность одиночного сокращения, иннервируемых ими мыш. волокон. |