Коллоквиум по возбудимым тканям. Вопросы коллоквиума по разделу Физиология возбудимых тканей
 Скачать 0.85 Mb. Скачать 0.85 Mb.
|

|
Тетаническое сокращение – длительное сокращение скелетных мыш. волокон. В его основе лежит суммации одиночных мыш. сокращений. При нанесении на мыш. волокно 2х быстро следующих друг за другом раздражений амплитуда сокращения будет больше, чем при одиночном раздражении. Сократительные эффекты, вызванные первым и 2ым раздражениями, складываются, происходит суммация (суперпозиция) сокращений, при этом скольжение нитей актина и миозина относительно друг друга увеличивается. Для реализации суммации в одиночном волоке важно, чтобы следующее раздражение наносилось после исчезновения ПД волокна, т.е. после рефрактерного периода, вызванного предыдущим стимулом. 25. Условия возникновения оптимума и пессимума частоты и силы раздражения (Н.Е. Введенский). При сравнительно низких частотах раздражения наступает зубчатый тетанус, при большой частоте – гладкий тетанус. При зубчатом тетанусе каждый последующий нервный импульс воздействует на начавшую расслабляться мышцу, при этом происходит неполная суммация сокращений. При гладком тетанусе, имеющем бо́льшую амплитуду, воздействие импульса происходит в конце периода укорочения, что приводит к полной суммации сокращений. Если ритм раздражения увеличить примерно до 10 - 15 раз в 1 сек, следующие друг за другом по нерву раздражающие сигналы сближаются во времени настолько, что каждый последующий стимул будет воздействовать на мышцу до того, как она успеет полностью расслабиться после предшествующего сокращения. Тогда в ответ на второе, третье и последующие раздражения мышца будет сокращаться, исходя не из полностью, а только частично расслабленного состояния. Это позволит записать неполный, или зубчатый, тетанус. П 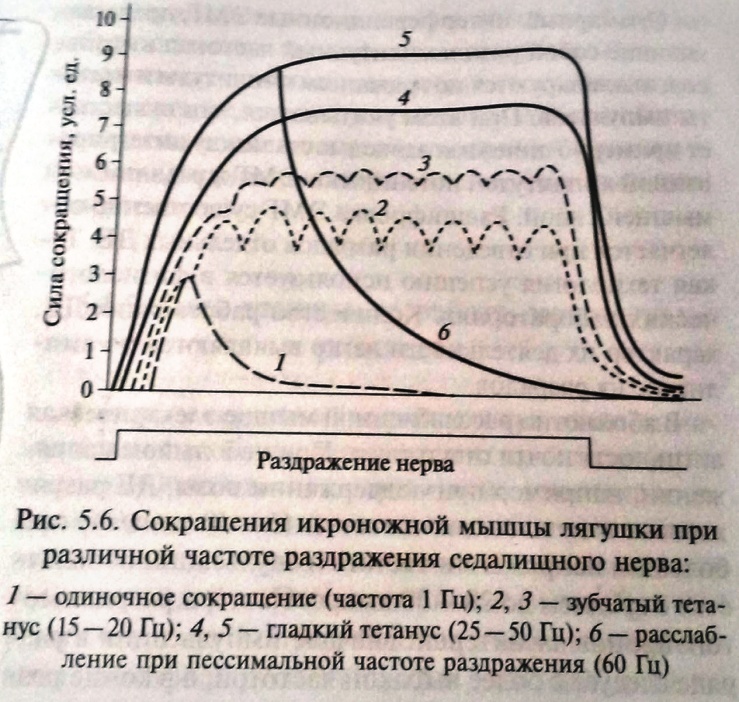 ри еще большем ритме раздражения (для икроножной мышцы лягушки не меньше 20 раз в 1 сек) следующие друг за другом импульсы сближаются в такой мере, что каждый последующий действует раньше, чем закончится восходящая часть предыдущего сокращения. И поскольку интервал между стимулами короче, чем фаза сокращения, расслабления в интервале между раздражениями вообще не происходит. Результатом является сплошной (гладкий) тетанус, характеризуемый длительным сокращением, не прерываемым расслаблениями. ри еще большем ритме раздражения (для икроножной мышцы лягушки не меньше 20 раз в 1 сек) следующие друг за другом импульсы сближаются в такой мере, что каждый последующий действует раньше, чем закончится восходящая часть предыдущего сокращения. И поскольку интервал между стимулами короче, чем фаза сокращения, расслабления в интервале между раздражениями вообще не происходит. Результатом является сплошной (гладкий) тетанус, характеризуемый длительным сокращением, не прерываемым расслаблениями. Оптимум и пессимум частоты раздражения. Н. Е. Введенский показал, что величина тетануса может значительно отличаться от величины одиночных сокращений, в зависимости от силы и частоты импульсов. Умеренные по силе и частоте раздражения (оптимальные), вызывают максимальный эффект, значительно превышающий амплитуду одиночного сокращения, а очень сильные и частые – (пессимальные), вызывают заметное ослабление эффекта. На основании этого Н. Е. Введенский сделал вывод, что высота тетануса определяется не только наложением отдельных сокращений друг на друга, но и теми функциональными изменениями, которые оставляют в ткани приходящие раздражения. Если каждый последующий импульс приходит с таким интервалом, что застает ткань в состоянии повышенной реактивной способности, то эффект сокращения будет сильным, а высота тетануса больше. Н. Е. Введенский назвал это состояние повышенной возбудимости вслед за протекшим сократительным эффектом экзальтационной фазой. Экзальтационной фазе предшествует состояние пониженной реактивной способности – рефрактерная фаза. Импульсы, следующие с такой частотой, при которой они попадают в рефрактерную фазу, вызывают пессимальный эффект, снижение высоты тетануса. Это явление называют пессимальным торможением. Таким образом, пессимальным или оптимальным будет ответ, зависит от того, в каком состоянии находится ткань к моменту прихода действующего на нее импульса. Для свежей, неутомленной икроножной мышцы лягушки оптимальная частота раздражений, дающая при физиологически максимальной силе раздражения тетанус наибольшей величины, составляет 50-100 раз в1 сек; увеличение частоты раздражения до 200 - 300 раз в 1 сек приводит к пессимальному эффекту. 26. Работа и сила мышц. Динамометрия и эргография. Теория утомления. Гипертрофия и атрофия мышц. Мышечная сила - мера механического воздействия на мышцу со стороны других тел, которая выражается в ньютонах или кг-силах. При изотоническом сокращении в эксперименте сила определяется массой максимального груза, который мышца может поднять (динамическая сила), при изометрическом - максимальным напряжением, которое она может развить (статическая сила). Сила зависит от: 1) ДЕ (быстрая, медленная). 2) Силы импульса. 3) Физиологических св-в мышцы. Мышечная сила при прочих равных условиях определяется обычно поперечным сечением мышцы. В некоторых мышцах (нпр, портняжной) все волокна параллельны их длинной оси – это параллельно-волокнистый тип мышцы. В мышцах перистого типа волокна расположены косо, с одной стороны они прикреплены к центральному сухожилию, а с другой – к центр. сухожильному футляру. Физиологическое сечение, т.е. сумма поперечных сечений всех волокон, совпадает с геометрическим только в мышцах с продольно расположенными волокнами, у мышц с косым расположением волокон первое может значительно превосходить второе. Чем больше физиологическое сечение, тем больше груз, который она в состоянии поднять. Для сравнения силы разных мышц делят максимальный груз, который они в состоянии поднять, на площадь их поперечного физиолг. сечения – удельная сила мышцы. 4) Силы растяжения – если перетянута – толстые и тонкие нити её саркомеров не перекрываются, общая сила мышцы равна 0; если натуральная величина – все головки миелиновых нитей способны контактировать с актиновыми нитями, сила мыш. скоращения возрастает до максимума. При дальнейшем уменьшении длины мыш. волокон из-за «заползания» тонких нитей с соседние саркомеры и уменьшения возможной хоны контакта нитей актина и миозина сила мыш. сокращения снова уменьшается. Работа мышцы измеряется произведением поднятого груза на величину её укорочения. Зависимость мыш. работы от нагрузки подчиняется закону средних нагрузок. Если мышца сокращается без нагрузки, её вешняя работа равна нулю. По мере увеличения груза работа увеличивается, достигая максимума средних нагрузок. Затем она постепенно уменьшается. Работа становится равной 0 при очень большом грузе, который мышца поднять не способна. Утомление мышцы выражается в уменьшении силы её сокращения, скорости укорочения и расслабления, в результате работа и мощность также уменьшаются. В эксперименте при длительном частом раздражении сила сокращений мышцы уменьшается, вплоть до полного отсутствия сокращений. Скорость развития утомления зависит от ритма работы и величины груза. В условиях целостного организма физическое утомление развивается и в ЦНС (центральное утомление) – в настоящее время это вторично. При физической работе увеличивается выброс в кровь адреналина и норадреналина, которые стимулируют работу Na/К-насоса. что тормозит развитие утомления. Напротив, серотонин ускоряет процессы центрального утомления; дофамин, тормозя синтез серотонина в нейронах ЦНС, задерживает развитие центрального утомления при физической нагрузке. Динамометрия - метод измерения силы. Эргография - метод изучения процессов утомления и других аспектов мышечной деятельности, основанный на графической регистрации частоты и амплитуды движений пальца или сгибания руки в локтевом суставе при подъеме груза на определенную высоту. Гипертрофия и атрофия мышц. Увеличение общей массы мышцы называют мышечной гипертрофией, а уменьшение — мышечной атрофией. Выделяют два вида гипертрофии: 1. Миофибриллярный тип. Развивается при статической работе (поднятие тяжести). При этом типе гипертрофии увеличивается число миофибрилл и значительно увел. сила мышцы. Например, тяжелоатлеты. 2. Саркоплазматический тип – увеличение объема саркоплазмы (гликогена, креатининфосфата, миоглобина, числа капилляров). При этом типе гипертрофии развивается выносливость. Например, бегуны на длинной дистанции. Мышечная гипертрофия практически всегда является результатом увеличения количества актиновых и миозиновых нитей в каждом мышечном волокне, что ведет к их укрупнению. Это называют простой гипертрофией волокон. Степень гипертрофии значительно возрастает, если во время сокращения мышца нагружена. Для развития значительной гипертрофии достаточно лишь нескольких сильных сокращений в день в течение 6-10 нед. Атрофия мышцы развивается при ее бездеятельности. Атрофия способствует постельный режим, перерезка сухожилий, заболевания нервной системы, гипсовая повязка. 27. Физиологические особенности и свойства гладких мышц, их значение в миогенной регуляции моторной функции внутренних органов. Гладкие мышцы построены из веретенообразных одноядерных мышечных клеток. Толщина 2-10 мкм, длина 50-400 мкм. Несмотря на наличие межклеточных щелей шириной 60-150 нм, гладкая мышца функционирует как синцитий – функциональное образование, в котором возбуждение способно беспрепятственно передаваться с одной клетки на другую по крайней мере в пределах одного мышечного пучка, являющегося обычно функциональной единицей гладкой мышцы – это унитарные гладкие мышцы. Мышечные же волокна бронхов и крупных артерий имеет мало щелевых контактов, поэтому сокращаются изолировано друг от друга. Такие гладкие мышцы называют мультиунитарными. Каждое волокно такой мышцы получает отдельную иннервацию. Гладкие мышцы имеют свои особенности: 1) нестабильный мембранный потенциал, который поддерживает мышцы в состоянии постоянного частичного сокращения – тонуса; 2) самопроизвольная автоматическая активность; 3) сокращение в ответ на растяжение; 4) пластичность (уменьшение растяжения при увеличении растяжения); 5) высокую чувствительность к химическим веществам. 6) медлительность сокращения. Латентный период сокращения гладкой мышцы в 300 раз превышает скрытый период скелетного. Продолжительность сокращения в желудке кролика достигает 5, а в желудке лягушки - 60 с и более. Миогенная регуляция. Тонус сосудов во многом определяет параметры системной гемодинамики и регулируется миогенными, гуморальными и нейрогенными механизмами. В основе миогенного механизма лежит способность гладких мышц сосудистой стенки возбуждаться при растяжении. Именно автоматия гладких мышц создает базальный тонус многих сосудов, поддерживают начальный уровень давления в сосудистой системе. В сосудах кожи, мышц, внутренних органов миогенная регуляция тонуса играет относительно небольшую роль. Но в почеченых, мозговых и коронарных ссудах она является ведущей и поддерживает нормальный кровоток в широком диапазоне артериального давления. 28. Особенности сокращения и передачи возбуждения в гладких мышцах. П 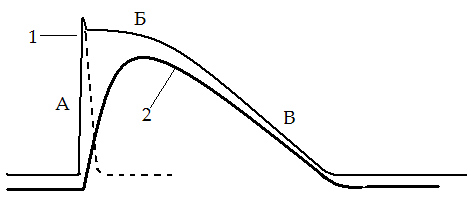 отенциал действия (1) и кривая сокращения (2) гладкомышечной клетки. отенциал действия (1) и кривая сокращения (2) гладкомышечной клетки. А – фаза деполяризации (Na+- вход); Б – «кальциевое плато» (Ca2+- вход); В – фаза реполяризации (К+- выход); (пунктиром ПД скел. мышцы) Особенности сокращения. Процесс сокращения гладкомышечных волокон совершается по тому же механизму скольжения нитей актина и миозина относительно друг друга, что и в скелетных мышцах, однако у гладкомышечных клеток нет той стройной оранжировки сократительных белков, как у скелетных мышц. У этих клеток миофибриллы с саркомерами расположены нерегулярно, поэтому клетка не имеет поперечной исчерченности. Электромеханическое сопряжение в гладкомышечных клетках идет иначе, чем в скелетных мышцах, так как в них слабо выражен саркоплазматический ретикулум. В связи с этим триггером для мышечного сокращения служит поступление ионов Са2+ в клетку из межклеточной среды в процессе генерации ПД. Того количества кальция, которое входит в клетку при возбуждении, вполне достаточно для полноценного фазного сокращения. Инициация сокращений гладких мышц с помощью ионов Са2+ также имеет несколько иной механизм, чем в п-п. волокнах. Ионы Са2+ воздействуют на белок кальмодулин, который активирует киназы легких цепей миозина. Это обеспечивает перенос фосфатной группы на миозин и сразу вызывает сокращение поперечных мостиков. О существовании тропонин-тропомиозиновой системы сведений нет. При снижении в миоплазме концентрации ионов Са2+ фосфатаза дефосфорилирует миозин и он перестает связываться с актином. Скорость сокращения гладких мышц невелика, она на 1 — 2 порядка ниже, чем у скелетных мышц. Сила сокращений некоторых гладких мышц позвоночных не уступает силе сокращений скелетных мышц. М 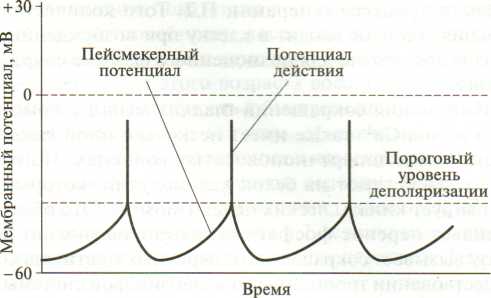 еханизм возбуждения гладкой мышцы. ПП некоторых гладкмыш. волокон, обладающих автоматией, обнаруживает постоянные небольшие колебания, обеспечивающие постоянные небольшие колебания, обеспечивающие возникновение ПД при достижении КП. Величина мембранного потенциала меньше значения мембранного потенциала меньше значения мембранного потенциала скелетных мышц и составляет в волокнах, не обладающих автоматией, - -60…-70 мВ, она несколько ниже в спонтанно активных клетках - -30…-70 мВ. еханизм возбуждения гладкой мышцы. ПП некоторых гладкмыш. волокон, обладающих автоматией, обнаруживает постоянные небольшие колебания, обеспечивающие постоянные небольшие колебания, обеспечивающие возникновение ПД при достижении КП. Величина мембранного потенциала меньше значения мембранного потенциала меньше значения мембранного потенциала скелетных мышц и составляет в волокнах, не обладающих автоматией, - -60…-70 мВ, она несколько ниже в спонтанно активных клетках - -30…-70 мВ. ПД. В гладких мышцах внутренних органов регистируются ПД 2х основных типов: пикоподобные ПД и ПД с выраженным плато. Длительность пикоподобных плато ПД составляет 5-80 мс, ПД с плато, характерное для гладких мышц мптки, уретры и некоторых сосудов, длятся 30-500 мс. Ионный механизм возникновения ПД в гладких мышцах: при удалении из омывающего раствора ионов Na+ (замена их ионами Li+ или холина) возникают полноценные ПД у гладких мышц. Удаление из раствора ионов Са2+ или воздействие на мышечные клетки блокаторов кальциевых каналов (например, верапамила) ведет к обратимому угнетению ПД. Все эти факты свидетельствуют о главной роли ионов Са2+ в генерации ПД гладких мышц. Электровозбудимые медленные Са-каналы обладают меньшей ионной избирательностью, чем «быстрые» натриевые каналы нервных и поперечно-исчерченных мышечных волокон. Помимо двухвалентных катионов они проницаемы и для ионов Na+. ПД гладких мышц, состоящие из начального пикового компонента и последующего плато, имеют более сложную ионную природу. Например, в гладких мышцах мочеточников начальный фрагмент ПД имеет преимущественно кальциевую природу, а последующий медленный компонент (плато) — преимущественно натриевую природу. |