Коллоквиум по возбудимым тканям. Вопросы коллоквиума по разделу Физиология возбудимых тканей
 Скачать 0.85 Mb. Скачать 0.85 Mb.
|

|
Волокна А – афф. и эфф. волокна ПНС (соматические). Волокна В – преганглионарные волокна ВНС. Волокна С – постганглионарные волокна ВНС, афф. волокна от обонятельных, некоторых болевых, тепловых и висцерал. рецепторов. Средний размер диаметра волокна уменьш. от типа А до С. Соответственно этому снижается и скорость проведения возбуждения. Лабильность уменьш. от волокон А до С и находится в обратной зависимости от продолжительности фазы абсолютной рефрактерности. Возбудимость также уменьш. от А до С. К давлению наиболее чувствительны волокна А, к гипоксии- В, к местным анестетикам – С. Законы проведения возбуждения: 1) Двустороннее проведение возбуждения: если в эксперименте нанести раздражение в любом участке нерва или нервного волокна, то возбуждение регистрируется как в проксимальном, итак и дистальном участках нервах или волокна. 2) Изолированное проведение возбуждения в отдельных волокнах нервного ствола: эл. поле ПД каждого волока гасится межклеточной жидкостью и оболочкой нервных волокон. Если одновременно возбуждается значительное количество волокон, то возможны суммация электрических полей ПД и возбуждение соседних волокон, что усиливает нервное влияние. 3) Физиологическая и анатомическая целостность – возбуждение может распространяться по неврному волокну только в случае его морф. и функц. целостности. 4) Большая скорость проведения возбуждения достигает 130 м/с в нервных волокнах Аа. Большая скорость распространения ПД обеспечивает быстрое влияние на другие нейроны и рабочие органы, а также получение обратной информации. 5) Малая утомляемость нервного волокна. Это обусловлено тем, что при проведении ПД по нервным волокнам используется незначительная часть запасов трансмембранных ионных градиентов и, следовательно, нужны небольшие количества АТФ для их восстановления. 6) Возможность функционального блока проведения возбуждения при морфологической целостности волокон. При действии различных факторов на нерв, вызывающих длительную деполяризацию клеточной мембраны, возникает полный блок проведения нервных импульсов (состояние парабиоза). Для возникновения блока в проведении возбуждения протяженность парабиотического участка должна превысить 5 мм в толстом миелинизированном волокне (в безмиелиновом 0,1 —1,0 мм), иначе ПД может возбуждать соседний участок волокна действием своего электрического поля. Нарушение физиологической непрерывности нервных волокон возникает при действии анестетиков, гипоксии, при воспалении, охлаждении. 7) Высокая лабильность нервное волокно может проводить до 200 — 400 имп/с. 12. Механизмы проведения возбуждения в мякотных и безмякотных нервных волокнах. В процессе проведения ПД можно выделить 2 этапа: этап распространения эл. поля, снижающего мембранный потенциал, и этап генерации новых ПД в новых участках нервного волокна. 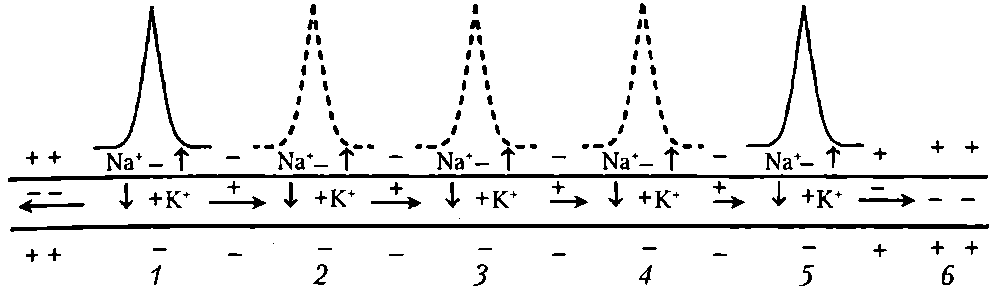 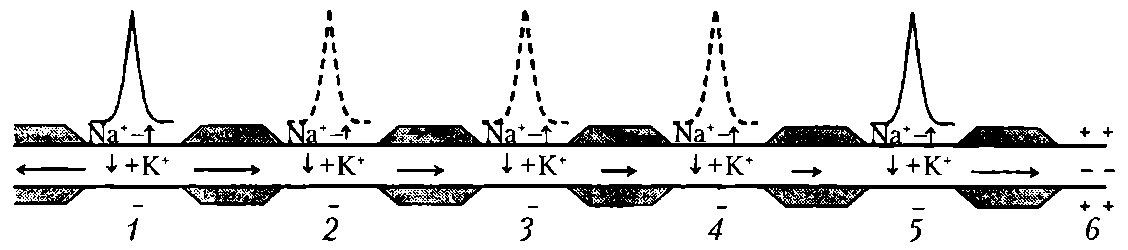 1) Непрерывное проведение ПД происходит в мышечных волокнах волокнах и в безмиелиновых нервных волокнах (тип С), имеющих равномерное распределение потенциалуправляемых ионных каналов по всей длине волокна, участвующих в генерации ПД. Проведение ПД начиначется с рапространения колеблющегося по величине эл. поля ПД. Амплитуда ПД в нерв. волокне (ПП+инверсия)= 100-120 мВ. Эл. поле ПД немиел. волокна может деполяризовать мембрану до КП на расстояние 0,1-1,0 мм. Это означает, что на этом участке одновременно генерируются новые ПД, обусловленные движением Na+ в клетку, K+ - из. Число одновременно возникающих ПД ограничивается длиной возбужденного участка. Для безмиел. волокна 0,1-1,0 мм, причём ПД возникают рядом друг с другом. Сами ПД не перемещ. (исчезли. где возникли). Главную роль в возникновении новых ПД играет передний ПД. Вспомогат. роль в генерации новых ПД играют промежуточные ПД (позади переднего) – их эл. поле суммируется с эл. полем переднего ПД, но они находятся дальше от участка нервного волокна, где возникают новые ПД. Т.е. возникший в нервном волокне ПД обеспечивает за счёт своего эл. поля возникновение новых ПД в соседнем участке нервного волокна и так далее – до окончания волокна («эстафета»). 2) Сальтоторное (скачкообразное) проведение ПД характерно для миел. волокон. Для этих волокон характерно конц. потенциалуправляемых ионных каналов только в перехватах Ранвье. В области миелиновых муфт, обладающих хорошими изолирующими св-вами, потенциалуправляемых каналов почти нет, поэтому ПД здесь не возникают. Покрытый миелиновой муфтой участок нервного волокна между перехватами Ранвье в механизме проведения ПД выполняет роль изолятора. В этих условиях ПД, возникший в одном перехвате Ранвье, за счёт своего эл. поля деполяризует мембрану соседних перехватов до КП на расстоянии до 5 мм, что приводит к возникновению в них новых ПД, т.е. возбуждение возникает как бы скачкообразно – только в перехватах. Благодаря этому в случае повреждения ближайших перехватов Ранвье на пути следования ПД он возбуждает 2-4-й и даже 5-й перехваты. Поэтому возбуждение распространяется очень быстро по всей длине волокна, а ионы движутся только перпендикулярно относительно длины волокна – внутрь волокон и их него (вдоль не успевают). Эл. поле ПД, возникших позади переднего, сумм. с эл. полем переднего ПД, как и при непрерывном распространении возбуждения. 2 преимущества сальтоторного проведения: 1) более экономично в энерг. плане, т.к. возбуждаются только перехваты Ранвье, площадь которых менее 1% мембраны, и поэтому меньше энергии для восстановления трансмембранных градиентов Nф+ и К+., расходующихся в процессе возникновения ПД. 2) сальтоторное проведение в миел. волокнах проводится с большей скоростью, чем в безмиел. волокнах, т.к. в них эл. поле ПД расп. значительно дальше – до 5мм (в безмиел. до 1 мм) на соседние перехваты Ранвье, поскольку электроизоляция уменьшается рассеивание эл. поля. 13. Трофическая функция двигательных нервных волокон. Нервные волокна имеют две основных функции - проведение возбуждения и аксонный транспорт, выполняющий трофическую функцию. Транспорт в отростках нейрона лучше изучен в аксонах и получил название аксонного транспорта. С помощью этого процесса осуществляется трофическое влияние не только в пределах различных участков нейрона, но и на иннервируемые клетки. Известны два вида аксонного транспорта — быстрый и медленный. Быстрый аксонный транспорт идет от тела клетки до аксонных окончаний (прямой транспорт, 250-400 мм/сут) и в противоположном направлении (обратный, или ретроградный. транспорт, 200-300 мм/сут). Посредством прямого транспорта в аксонные окончания доставляются ферменты, медиаторы, липиды, везикулы, содержащие гликопротеины мембран, специальные белки и пептиды (нейротрофогены). Посредством обратного транспорта в тело нейрона переносятся везикулы, содержащие остатки разрушенных структур, фрагментов и другие ростовые факторы, регулирующие синтез белка в соме клетки. Многие вещества, доставленные путем ретроградного транспорта, подвергаются разрушению в лизосомах. Быстрый аксонный транспорт осуществляется с помощью микротрубочек и микрофиламентов, часть которых представляет собой актиновые нити (актин составляет 10—15% белков нейрона). Для транспорта необходима энергия АТФ. Снижение уровня АТФ в аксоне более чем в 2 раза, падение концентрации Са2+, разрушение микротрубочек (например, колхицином) и микрофиломентов (цитохолазином В), блокирует аксонный транспорт . Медленный аксонный транспорт осущ. только в прямом направлении и представляет собой передвижение всего стоба аксоплазмы. Скорость медленнпгп транспорта равна 1—2 мм/сут, что соответствует скорости роста аксона в онтогенезе и при регенерации. С помощью этого транспорта перемещаются образованные в ЭПС белки микротрубочек и микрофиламентов (тубулин, актин и др.), ферменты цитозоля, РНК, белки каналов, насосов и другие вещества. Механизмы медленного и быстрого аксонного транспорта различны: медленный транспорт не нарушается при разрушении микротрубочек, но прекращается при отделении аксона от тела нейрона. Прямой и обратный транспорт белков и други веществ необходим для поддержания структуры функции аксона и его пресинаптических окончание а также для таких процессов, как аксонный рост образование синаптических контактов, т. е. аксонный транспорт выполняет внутриклеточную трофическую роль. Аксонный транспорт играет важную роль при регенерации нервных волокон. Аксонный транспорт необходим для поддержания структуры нервного волокна. 14. Виды передач сигнала между возбудимыми клетками. Понятие синапса, классиф. синапсов. С 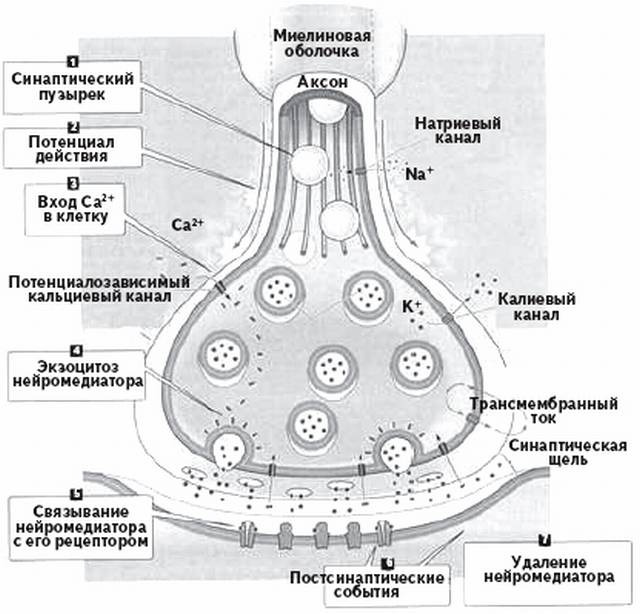 инапс – это специализированная структура, обеспечивающая передачу сигнала от клетки к клетке с помощью химических вещ-в или эл. поля. В составе синапса входит пре- и постсинаптическая мембраны и синаптическая щель. В области пресинаптической мембраны есть многочисленные везикулы, диаметром от 50 до 60 нм. Каждая синаптическая везикула содержит от 6000 до 10000 молекул медиатора ацетилхолина. Синаптические везикулы концентрируются в специализированных областях пресинаптической мембраны, которые наз. активными зонами. Постсинаптическая мембрана образует постсинаптические складки, которые увеличивают площадь мембраны. инапс – это специализированная структура, обеспечивающая передачу сигнала от клетки к клетке с помощью химических вещ-в или эл. поля. В составе синапса входит пре- и постсинаптическая мембраны и синаптическая щель. В области пресинаптической мембраны есть многочисленные везикулы, диаметром от 50 до 60 нм. Каждая синаптическая везикула содержит от 6000 до 10000 молекул медиатора ацетилхолина. Синаптические везикулы концентрируются в специализированных областях пресинаптической мембраны, которые наз. активными зонами. Постсинаптическая мембрана образует постсинаптические складки, которые увеличивают площадь мембраны.Синаптическая щель (50 нм) заполнена межклеточной жидкостью, а также разнообразными элементами межклеточного матрикса. Здесь есть базальная пластинка, которая содержит высокие конц. АХЭ, быстро разрушающей высвобожденный ацетилхолин и готовящая синапс к проведению очередного сигнала. 1) По морфологическому принципу (виды передач сигнала ): • нейро-мышечные (аксон нейрона контактирует с мышечной клеткой); • нейро-секреторные (аксон нейрона контактирует с секреторной клеткой); • нейро-нейрональные (аксон нейрона контактирует с другим нейроном): • аксо-соматические (с телом другого нейрона), • аксо-аксональные (с аксоном другого нейрона), • аксо-дендритические (с дендритом другого нейрон). 2) По способу передачи возбуждения: • электрические (возбуждение передается при помощи электрического тока); • химические (возбуждение передается при помощи химического вещества): •адренергические (возбуждение передается при помощи норадреналина), •холинергические (возбуждение передается при помощи ацетилхолина), •пептидергические, NO -ергические, пуринергические и т. п. 3) По физиологическому эффекту: • возбуждающие (деполяризуют постсинапт. мембрану и вызывают возбуждение постсинапт.клетки); • тормозные (гиперполяризуют постсинапт. мембрану и вызывают торможение постсинапт. клетки). 15. Функциональные свойства эл. и хим. синапсов. Механизм формирования ВПСП, ТПСП. Одностороннее проведение возбуждения в химическом синапсе связано с его функциональной асимметрией: молекулы медиатора выделяются только на пресинаптической мембране, а рецепторы медиатора расположены только на постсинаптической мембране. Высокая утомляемость химического синапса объясняется истощением запасов медиатора. Утомляемость электрического синапса соответствует утомляемости нервного волокна. Низкая лабильность химического синапса определяется главным образом периодом рефрактерности хемочувствительных каналов на постсинаптической мембране.
Чувствительность синапса к внешним воздействиям определяется характером процессов, протекающих в синапсе при передаче возбуждения. Химические синапсы чувствительны к действию химических веществ, влияющих на синтез и секрецию медиатора, взаимодействие медиатора с рецептором. По знаку действия: возбуждающие и тормозные. Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембраны, которая может вызвать потенциал действия при определённых условиях.), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Тормозные синапсы бывают двух видов: 1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала; 2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение. Тормозной синапс. Нервный импульс, идущий по волокну, достигает пресинаптической мембраны синаптической бляшки, изменяя ее проницаемость для ионов кальция. Ионы кальция поступают внутрь бляшки, и под их воздействием пузырьки с тормозным медиатором подходят к внутренней поверхности пресинаптической мембраны, лопаются, и медиатор, изливаясь в синаптическую щель, диффундирует через нее и действует на рецепторы постсинаптической мембраны, увеличивая ее проницаемость для ионов калия. Т.к. ионов калия больше внутри, он выходит из клетки; увеличивается ПП, развивается гиперполяризация и тормозной постсинаптический потенциал (ТПСП), наступает торможение. Возбуждающий синапс. Нервный импульс, идущий по волокну, достигает пресинаптической мембраны синаптической бляшки, изменяя ее проницаемость для ионов кальция. Ионы кальция поступают внутрь бляшки, и под их воздействием пузырьки с возбуждающим медиатором подходят к внутренней поверхности пресинаптической мембраны, лопаются, и медиатор, изливаясь в синаптическую щель, диффундирует через нее и действует на рецепторы постсинаптической мембраны, увеличивая ее проницаемость для ионов натрия. Т.к. ионов натрия больше снаружи, он входит внутрь клетки и обуславливает деполяризацию мембраны; возникает ПД и возбуждающий постсинаптический потенциал (ВПСП), клетка возбуждается. 16. Характеристика мионеврал. синапса. Механизм передачи возбуждения с нерва на мышцу. 17. Потенциал концевой пластинки (ПКП), миниатюрные потенциалы концевой пластинки (МПКП), их физико-химическая природа, параметры, свойства и функциональное значение. Процесс передачи сигнала с нервного волокна на мышечное включает 4 основных этапа. 1) Выброс медиатора в синаптическую щель. Он запускается посредством ПД пресинапт. окончания – около 100 квантов медиаторов на один ПД. В двиг. окончании этого кол-ва достаточно для проведения 10 000 импульсов. Деполяризация мембраны пресинапт. окончания ведёт к открытию потенциалуправляемых Са-каналов. Са2+ входит в нервное окончание согласно электрохим. градиенту. Часть медиатора в пресинапт. окончании локализуется на пресинапт. мембране изнутри. Са2+ активирует экзоцитозный аппарат пресинапса, представляющий собой совокупность белков пресинапт. окончания. Это обеспечивает выброс АХ посредством экзоцитоза в синапт. щель. Кол-во высвобождаемого АХ из пресинапт. окончания пропорц в 4-ой степени кол-ву вступившего туда Са2+. 2) Диффузия АХ к постасинапт. мембране, где локализуются N-холинрецепторы. Длится 1 мс. 3) Возникновение возбуждения в мыш. волокне. В результате взаимодействия АХ с N-холинорецепторами постсинапт. мембраны открывается ионные канале на 1 мс и вследствие преобладания входа Na+ в клетку происходит деполяризация постсинапт. мембраны (мембраны концевой пластинки). Эту деполяризацию в неврно-мыш. синапсе наз. потенциалом концевой пластинки (ПКП). Особенностью нервно-мышечного синапса скелетного мышечного волокна является то, что при одиночной его активации формируется ПКП большой амплитуды (30-40 мВ), и его эл. поле достаточно для генерации ПД на мембране мыш. волокна вблизи синапса. Большая амплитуда ПКП связана с тем, что нервные окончания делятся на много веточек, каждая из которых выбрасывает медиатор. |