Вопросы к занятию №2. Возбудимые ткани
 Скачать 417.93 Kb. Скачать 417.93 Kb.
|

|
ТЕМА: ВОЗБУДИМЫЕ ТКАНИ Структурно-функциональная организация мышечного волокна. Физиологические свойства скелетной мышцы. Структурной и функциональной единицей скелетной мышцы является мышечное волокно, представляющее собой вытянутую (длиной 0,5—40 см) многоядерную клетку. Толщина мышечных волокон составляет 10— 100 мкм. Диаметр их может увеличиваться при интенсивных тренировочных нагрузках, количество же мышечных волокон может нарастать лишь до 3—4-месячного возраста. Мембрану мышечного волокна называютсарколеммой, цитоплазму —саркоплазмой. Сарколемма. Сарколеммой называют клеточную мембрану мышечного волокна. Она состоит из собственно мембраны клетки, называемой плазматической мембраной, и наружного покрытия из тонкого слоя полисахаридного материала, который содержит множество тонких коллагеновых нитей. На концах мышечного волокна этот поверхностный слой сарколеммы сливается с сухожильными волокнами. Сухожильные волокна, в свою очередь, собираются в пучки и формируют мышечные сухожилия, вплетающиеся в кости. Миофибриллы. Нити актина и миозина. Каждое мышечное волокно содержит от нескольких сотен до нескольких тысяч миофибрилл, которые представлены на поперечном срезе в виде небольших светлых точек. Каждая миофибрилла собрана примерно из 1500 смежных нитей миозина и 3000 нитей актина, представляющих собой большие полимеризованные белковые молекулы, ответственные за мышечное сокращение. Актиновые и миозиновые нити частично перекрываются, в связи с чем миофибриллы имеют чередующиеся светлые и темные полоски, если смотреть на электронную фотографию. Концы актиновых нитей прикреплены к так называемому Z-диску (Z-мембране). Отсюда актиновые нити распространяются в обе стороны, проходя между миозиновыми нитями. Z-диск, состоящий из нитевидных белков, проходит в поперечном направлении через миофибриллу, а также через все мышечное волокно, переходя от одной миофибриллы к другой, скрепляя их между собой. Часть миофибриллы (или целого мышечного волокна), расположенную между двумя последовательными Z-дисками, называют саркомером. В сокращенном мышечном волокне длина саркомера равна примерно 2 мкм. При этой длине актиновые нити полностью перекрывают миозиновые, и кончики актиновых нитей начинают перекрывать друг друга. Именно при этой длине мышца способна генерировать максимальную силу сокращения. Удерживать актиновые и миозиновые нити рядом довольно трудно. Это обеспечивается с помощью большого числа нитевидных молекул белка, называемого тайтином. Упругие молекулы тайтина образуют каркас, удерживающий актиновые и миозиновые нити в положении, обеспечивающем нормальную работу сократительного аппарата саркомера В поперечном сечении мышцы также можно увидеть Т-трубочки (стрелки), которые связаны с внеклеточным пространством и отвечают за проведение электрического сигнала в центр мышечного волокна. Саркоплазма. В каждом мышечном волокне многочисленные миофибриллы расположены рядом, бок о бок. Пространство между ними заполнено внутриклеточной жидкостью, называемой саркоплазмой и содержащей большое количество калия, магния и фосфатов, а также множество ферментов. Кроме того, имеется огромное число митохондрий, расположенных параллельно миофибриллам. Они снабжают сокращающиеся миофибриллы большим количеством энергии в форме аденозинтрифосфата, синтезируемого митохондриями. Саркоплазматический ретикулум. В саркоплазме, окружающей миофибриллы каждого мышечного волокна, имеется также хорошо развитый ретикулум, называемый саркоплазматическим ретикулумом. Он имеет специфическую организацию, исключительно важную для регуляции мышечного сокращения. Саркоплазматический ретикулум особенно сильно развит в очень быстро сокращающихся типах мышечных волокон (к белым мышечным волокнам). Механизм сопряжения возбуждения, сокращения и расслабления скелетной мышцы (электромеханическое сопряжение). Мышечное сокращение является жизненно важной функцией организма, связанной с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами. Все виды произвольных движений – ходьба, мимика, движения глазных яблок, глотание, дыхание и т. п. осуществляются за счет скелетных мышц. Непроизвольные движения (кроме сокращения сердца) – перистальтика желудка и кишечника, изменение тонуса кровеносных сосудов, поддержание тонуса мочевого пузыря – обусловлены сокращением гладких мышц. Работа сердца обеспечивается сокращением сердечной мускулатуры. Молекулярные механизмы сокращения скелетной мышцы Согласно теории скольжения нитей, мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых филламентов друг относительно друга. Механизм скольжения нитей включает несколько последовательных событий: Головки миозина присоединяются к центрам связывания актинового филламента. Взаимодействие миозина с актином приводит к конформационным перестройкам молекулы миозина. Головки приобретают АТФазную активность и поворачиваются на 120 °. За счет поворота головок нити актина и миозина передвигаются на «один шаг» друг относительно друга. Рассоединение актина и миозина и восстановление конформации головки происходит в результате присоединения к головке миозина молекулы АТФ и ее гидролиза в присутствии Са++. Цикл «связывание – изменение конформации – рассоединение – восстановление конформации» происходит много раз, в результате чего актиновые и миозиновые филламенты смещаются друг относительно друга, Z -диски саркомеров сближаются, и миофибрилла укорачивается. Фазы и режимы сокращения скелетной мышцы Фазы мышечного сокращения При раздражении скелетной мышцы одиночным импульсом электрического тока сверхпороговой силы возникает одиночное мышечное сокращение, в котором различают 3 фазы: латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения; возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия; фаза укорочения (около 50 мс); фаза расслабления (около 50 мс). Режимы мышечного сокращения В естественных условиях в организме одиночного мышечного сокращения не наблюдается, так как по двигательным нервам, иннервирующим мышцу, идут серии потенциалов действия. В зависимости от частоты приходящих к мышце нервных импульсов мышца может сокращаться в одном из трех режимов: Одиночные мышечные сокращения возникают при низкой частоте электрических импульсов. Если очередной импульс приходит в мышцу после завершения фазы расслабления, возникает серия последовательных одиночных сокращений. При более высокой частоте импульсов очередной импульс может совпасть с фазой расслабления предыдущего цикла сокращения. Амплитуда сокращений будет суммироваться, возникнет зубчатый тетанус – длительное сокращение, прерываемое периодами неполного расслабления мышцы. При дальнейшем увеличении частоты импульсов каждый следующий импульс будет действовать на мышцу во время фазы укорочения, в результате чего возникнет гладкий тетанус – длительное сокращение, не прерываемое периодами расслабления. Оптимум и пессимум частоты Амплитуда тетанического сокращения зависит от частоты импульсов, раздражающих мышцу. Оптимумом частоты называют такую частоту раздражающих импульсов, при которой каждый последующий импульс совпадает с фазой повышенной возбудимости и соответственно вызывает тетанус наибольшей амплитуды. Пессимумом частоты называют более высокую частоту раздражения, при которой каждый последующий импульс тока попадает в фазу рефрактерности, в результате чего амплитуда тетануса значительно уменьшается. Работа скелетной мышцы Сила сокращения скелетной мышцы определяется 2 факторами: числом ДЕ (двигательная единица), участвующих в сокращении; частотой сокращения мышечных волокон. Работа скелетной мышцы совершается за счет согласованного изменения тонуса (напряжения) и длины мышцы во время сокращения. Виды работы скелетной мышцы: динамическая преодолевающая работа совершается, когда мышца, сокращаясь, перемещает тело или его части в пространстве; статическая (удерживающая) работа выполняется, если благодаря сокращению мышцы части тела сохраняются в определенном положении; динамическая уступающая работа совершается, если мышца функционирует, но при этом растягивается, так как совершаемого ею усилия недостаточно, чтобы переместить или удержать части тела. Во время выполнения работы мышца может сокращаться: изотонически – мышца укорачивается при постоянном напряжении (внешней нагрузке); изотоническое сокращение воспроизводится только в эксперименте; изометричеки – напряжение мышцы возрастает, а ее длина не изменяется; мышца сокращается изометрически при совершении статической работы; ауксотонически – напряжение мышцы изменяется по мере ее укорочения; ауксотоническое сокращение выполняется при динамической преодолевающей работе. Правило средних нагрузок – мышца может совершить максимальную работу при средних нагрузках. Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений, удлинением латентного периода сокращения и фазы расслабления. Причинами утомления являются: истощение запаса АТФ, накопление в мышце продуктов метаболизма. Утомляемость мышцы при ритмической работе меньше, чем утомляемость синапсов. Поэтому при совершении организмом мышечной работы утомление первоначально развивается на уровне синапсов ЦНС и нейромышечных синапсов. Расслабление скелетной мышцы Расслабление мышцы вызывается обратным переносом ионов Са++ посредством кальциевого насоса в каналы саркоплазматического ретикулума. По мере удаления Са++ из цитоплазмы открытых центров связывания становится все меньше и в конце концов актиновые и миозиновые филламенты полностью рассоединяются; наступает расслабление мышцы. Контрактурой называют стойкое длительное сокращение мышцы, сохраняющееся после прекращения действия раздражителя. Кратковременная контрактура может развиваться после тетанического сокращения в результате накопления в саркоплазме большого количества Са++; длительная (иногда необратимая) контрактура может возникать в результате отравления ядами, нарушений метаболизма. Электромеханическое сопряжение в миоците скелетной мышцы. Совокупность явлений, обусловливающих связь между возбуждением (ПД) и сокращением мышечных волокон – электромеханическое сопряжение. К регулярно расположенным Т‑трубочкам с двух сторон подходят терминальные цистерны саркоплазматического ретикулума (САР). Т‑трубочка и расположенные с двух сторон от неё цистерны образуют триаду. Z‑мембраны и зоны перекрытия актиновых и миозиновых нитей расположены рядом с триадами. Такая структура необходима для эффективного электромеханического сопряжения. Электромеханическое сопряжение включает следующие процессы: образование потенциала концевой пластинки (ПКП); электротоническое распространение ПКП в околосинаптическую область; генерация потенциала действия мышечного волокна в околосинаптической области; распространение потенциала действия по поверхностной мембране мышечного волокна; распространение потенциала действия по мембранам поперечных трубочек (Т‑системы); реакция рецепторов дигидропиридина на изменение мембранного потенциала; передача сигнала дигидропиридиновыми рецепторами рианодиновым рецепторам терминальных цистерн саркоплазматического ретикулума; открытие каналов Ca2+ с рианодиновыми рецепторами и выход Ca2+ из саркоплазматического ретикулума (цистерн) в саркоплазму (цитозоль); диффузия Ca2+ в межфибриллярное пространство; связывание Ca2+ с тропонином, расположенным на тонком, актиновом, миофиламенте; конформационные изменения тропонина при связывании Са2+ => «заталкивание» тропомиозина в канавки актиновых нитей. Соотношение кривых возбудимости, возбуждения и сокращения скелетной мышцы. Сопряжение возбуждения и сокращения в скелетной мышце В состоянии покоя скольжения нитей в миофибрилле не происходит, так как центры связывания на поверхности актина закрыты молекулами белка тропомиозина. Возбуждение (деполяризация) миофибриллы и собственно мышечное сокращение связаны с процессом элетромеханического сопряжения, который включает ряд последовательных событий. • В результате срабатывания нейромышечного синапса на постсинаптической мембране возникает ВПСП, который генерирует развитие потенциала действия в области, окружающей постсинаптическую мембрану. • Возбуждение (потенциал действия) распространяется по мембране миофибриллы и за счет системы поперечных трубочек достигает саркоплазматического ретикулума. Деполяризации мембраны саркоплазматического ретикулума приводит к открытию в ней Са++ -каналов, через которые в саркоплазму выходят ионы Са++. • Ионы Са++ связываются с белком тропонином. Тропонин изменяет свою конформацию и смещает молекулы белка тропомиозина, которые закрывали центры связывания актина. • К открывшимся центрам связывания присоединяются головки миозина, и начинается процесс сокращения. Для развития указанных процессов требуется некоторый период времени (10–20 мс). Время от момента возбуждения мышечного волокна (мышцы) до начала ее сокращения называют латентным периодом сокращения. ……………………………….......... При одиночном раздражении нерва мышца отвечает одиночным сокращением (рис. 16, А). В нем различают следующие фазы: латентный период - время от начала раздражения до начала укорочения мышцы. Длительность его 10-20 мс; он связан, прежде всего с инерционностью мышцы, биофизическими изменениями в ней; период укорочения (сокращения) возникает после латентного периода и длится до максимума сокращения (рис. 16, А); период расслабления - от максимума сокращения до возвращения мышцы в начальное состояние; длится несколько дольше периода сокращения. Общая длительность сокращения у мелких мышц, например, глазного яблока -- порядка 10 мс; икроножной - около 50 мс. При увеличении частоты сокращений происходит суммация одиночных сокращений, в результате чего возрастает их амплитуда сокращения (рис. 16, Б) и меняется характер сокращения. Зубчатый тетанус возникает в том случае, если каждое последующее раздражение попадает в период расслабления, а гладкий тетанус - в период сокращения (рис. 16, В). В организме в скелетных мышцах преобладают гладкотетанические сокращения, т.к. мотонейроны постоянно подают на мышцу нервные импульсы достаточно высокой частоты. Во время сокращения скелетная мышца изменяет свою возбудимость. Это изменение возбудимости рассматривают на примере различных мышц - (рис. 17) – скелетной мышцы, мышцы сердца и гладкой мышцы. 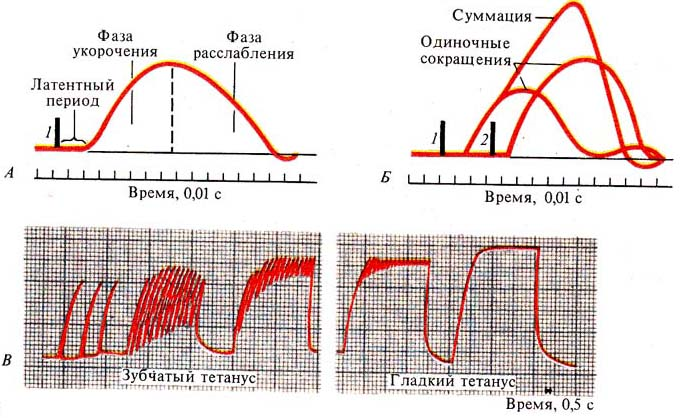 Рис.16. Одиночное сокращение (А), суммация одиночных сокращений (Б), тетанические сокращения (запись на кимографе) - (В): На рис. Б: I — момент первого раздражения, 2 — момент второго раздражения Наиболее короткой рефрактерностью обладает скелетная мышца, более длительный рефрактерный период - в мышце сердца; самый длительный – у гладкой мышцы (рис. 17). Для скелетной мышцы и мышцы сердца характерно наличие периода супернормальной возбудимости (рис. 17). Физиологическая роль рефрактерных периодов в мышцах – не допустить прерывания физиологического процесса сокращения случайным нервным импульсом; особенно это характерно для сердечной мышцы. 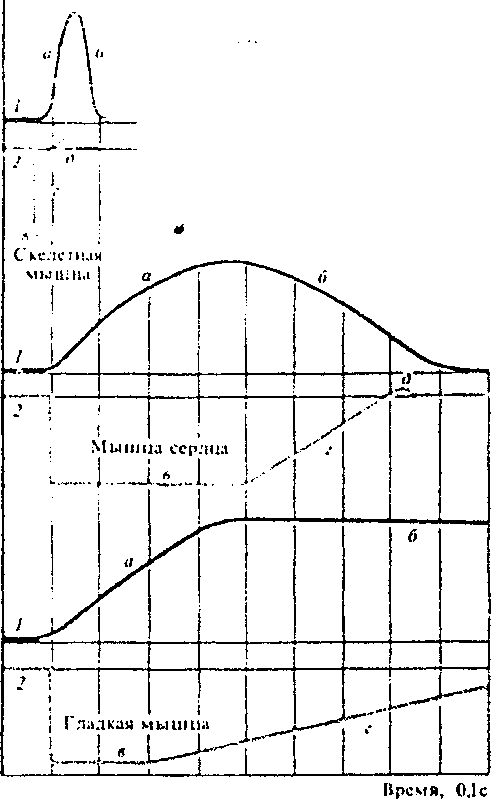 Рис.17. Взаимосвязь между сокращением и возбуждением мышц разного вида. 1 — кривая сокращения; 2 — кривая возбудимости; а — период сокращения, б — период расслабления; в — абсолютный рефрактерный период, г - относительный рефрактерный период; д - фаза супернормальной возбудимости. Определенную роль играет и повышение возбудимости скелетной мышцы - оно участвует в формировании оптимума сокращения при гладкотетанических сокращениях. Действие постоянного тока на возбудимые ткани. Полярный закон раздражения При раздражении нерва или мышцы постоянным током возбуждение возникает в момент замыкания постоянного тока только под катодом, а в момент размыкания - только под анодом. Эти факты объединяют под названием полярного закона раздражения Пфлюгера. Полярный закон доказывается следующими опытами: Умерщвляют участок нерва под одним из электродов, а второй электрод устанавливают на неповрежденном участке. Если с неповрежденным участком соприкасается катод, возбуждение возникает в момент замыкания тока: если же катод устанавливают на поврежденном участке, а анод - на неповрежденном, возбуждение возникает только при размыкании тока. Порог раздражения при размыкании, когда возбуждение возникает под анодом, значительно выше, чем при замыкании, когда возбуждение возникает под катодом. Изучение механизма полярного действия электрического тока стало возможным только после того, как был разработан описанный метод одновременного введения в тетки двух микроэлектродов: одного - для раздражения, другого- для отведения потенциалов. Было установлено, что потенциал действия возникает только в том случае, если катод находится снаружи, а анод- внутри клетки. При обратном расположении полюсов, т. е. наружном аноде и внутреннем катоде, возбуждения при замыкании тока не возникает, как бы силен он ни был. Прохождение через нервное или мышечное волокно электрического тока прежде всего вызывает изменения мембранного потенциал. В области приложения к поверхности ткани анода положительный потенциал на наружной стороне мембраны возрастает, т. е. происходит гиперполяризация, а в том случае, когда к поверхности приложен катод, положительный потенциал на наружной стороне мембраны снижается - возникает деполяризация. Как при замыкании, так и при размыкании тока изменения мембранного потенциала нервного- волокна не возникают и не исчезают мгновенно, а плавно развиваются во времени. Объясняется это тем, что поверхностная мембрана живой клетки обладает свойствами конденсатора. Обкладками этого «тканевого конденсатора» служат наружная и внутренняя поверхности мембраны, а диэлектриком - слой липидов, обладающий значительным сопротивлением. Ввиду наличия в мембране каналов, через которые могут проходить ионы, сопротивление этого слоя не равно бесконечности, как в идеальном конденсаторе. Поэтому поверхностную мембрану клетки обычно уподобляют конденсатору с параллельно включенным сопротивлением, по которому может происходить утечка зарядов. Временной ход изменений мембранного потенциала при включении и выключении тока зависит от емкости С и сопротивления мембраны R. Чем меньше произведение ДС- постоянная времени мембраны, тем быстрее при данной силе тока нарастает потенциал и, наоборот, большей величине соответствует меньшая скорость увеличения потенциала. Изменения мембранного потенциала возникают не только непосредственно в точках приложения к нервному волокну катода и анода постоянного тока, но и на некотором расстоянии от полюсов с той, однако, разницей, что их величина постепенно убывает по мере удаления от катода и анода. Объясняется это так называемыми кабельными свойствами нервного и мышечного волокон. Однородное нервное волокно в электрическом отношении представляет собой кабель, т. е. сердечник с низким удельным сопротивлением (аксоплазма)", покрытый изоляцией (мембраной), и помещенный в хорошо проводящую среду. При про г пускании, через некоторую точку волокна длительное время постоянного тока наблюдается стационарное состояние, при котором плотность тока и, следовательно, изменение мембранного" потенциала максимальны в месте приложения тока (т. е. непосредственно под катодом и анодом); с удалением от полюсов плотность тока и изменения потенциала на мембране экспоненциально уменьшаются по длине волокна. Поскольку рассматриваемые изменения мембранного потенциала в отличие от локального, ответа потенциала действия или следовых потенциалов не связаны с изменениями ионной проницаемости мембраны (т.-е. активным ответом волокна), их принято называть пассивными, или «электротоническими изменениями мембранного потенциала. В чистом виде последние могу быть зарегистрированы в условиях полной блокады ионных каналов химическими агентами. Различают кат- и ан-электротонические изменения потенциала, развивающиеся в области приложение соответственно катода и анода постоянного тока. Потенциал Критический уровень деполяризации - \ Регистрация изменений мембранного потенциала при внутриклеточном раздражении нервного или мышечного волокна показала, что потенциал действия возникает в. тот момент, когда деполяризация мембраны достигает критического уровня. Этот критический уровень деполяризации не зависит от характера примененного стимула, расстояния между электродами и т. п., а определяется исключительно свойствами самой мембраны. Скорость, с которой происходит деполяризация мембраны, при прочих равных 4 условиях зависит от силы раздражающего тока. При токе слабой силы деполяризация развивается медленно, поэтому. для возникновения потенциала действия стимул должен быть-большей длительности. В случае усиления раздражающего тока скорость развития деполяризации возрастает и. соответственно уменьшается минимальное время, необходимое для возникновения возбуждения. Чем быстрее развивается деполяризация мембраны, тем меньше минимальное время, необходимое для генерации потенциала действиями наоборот. Локальный ответ. В механизме критической деполяризации мембраны наряду с пассивными существенную роль играют активные подпороговые изменения мембранного потенциала, проявляющиеся й форме так называемого локального ответа. Виды и режимы сокращения скелетных мышц. Одиночное мышечное сокращение и его фазы. Тетанус и его виды. Оптимум и пессимум сокращения. Выделяют три виды мышечного сокращения: Одиночное мышечное сокращение; Тетаническое мышечное сокращение (тетанус); Тоническое мышечное сокращение. Кроме того, тетаническое мышечное сокращение делят на зубчатый и гладкий тетанус. Одиночное мышечное сокращение возникает в условиях действия на мышцу пороговых или надпороговых электрических стимулов, межимпульсный интервал которых равен или больше длительности одиночного мышечного сокращения. В одиночном мышечном сокращении выделяют три временных отрезка: латентный период, фазу укорочения и фазу расслабления. Самой короткой фазой является скрытый период, когда в мышце происходит электромеханическая передача. Фаза расслабления обычно в 1,5 – 2 раза более продолжительна, чем фаза сокращения, а при утомлении затягивается на значительное время. Тетаническое мышечное сокращение (тетанус) возникает в условиях действия на скелетную мышцу порогового или надпорогового электрического раздражителя, межимпульсный интервал которого меньше длительности одиночного мышечного сокращения. В зависимости от длительности межстимульных интервалов электрического раздражителя при его воздействии может возникнуть либо зубчатый, либо гладкий тетанус. Если межимпульсный интервал электрического раздражителя меньше длительности одиночного мышечного сокращения, но больше или равен сумме латентного периода и фазы укорочения, возникает зубчатый тетанус. Указанное условие выполняется при повышении частоты импульсного электрического раздражителя в определенном диапазоне. Если же длительность межимпульсного интервала электрического раздражителя меньше суммы латентного периода и фазы укорочения возникает гладкий тетанус. При этом амплитуда гладкого тетануса больше амплитуды и одиночного мышечного сокращения, и зубчатого тетанического сокращения. При дальнейшем уменьшении межимпульсного интервала электрического раздражителя, а, следовательно, при увеличении частоты, амплитуда тетанических сокращений возрастает. Однако, указанная закономерность не носит абсолютного характера: при определенном значении частоты вместо ожидаемого повышения амплитуды гладкого тетатнуса отмечается феномен ее снижения (см. рис. 5). Указанный феномен был впервые обнаружен Российским ученым Н.Е. Введенским и был назван пессимумом. В основе пессимальных явлений, по мнению Н.Е. Введенского лежит механизм торможения. Выделяют три режима мышечного сокращения: Изотонический. Изотонический режим мышечного сокращения характеризуется преимущественным изменением длины мышечного волокна, без существенного изменения напряжения. Указанный режим мышечного сокращения наблюдается, например, при поднятии легких и средних по массе грузов. Изометрический. Изометрический режим мышечного сокращения характеризуется преимущественным изменением мышечного напряжения, без существенного изменения длины. Примером может служить изменения состояния мышц при попытке человека сдвинуть с места предмет большой массы (например, при попытке сдвинуть с места стену в комнате). Смешанный (ауксометрический). Смешанный (ауксометрический) тип мышечного сокращения, наиболее реальный, наиболее часто встречающийся вариант. Содержит в себе компоненты первого и второго вариантов в разных соотношениях в зависимости от реальных условий окружающей среды. Н.Е. Введенский открыл явление оптимума и пессимума частоты раздражений (рис 3). На классическом нервно-мышечном препарате было установлено: при повышении частоты раздражений от 10 до 50 им/с увеличивается амплитуда мышечного сокращения; дальнейшее повышение частоты вызывает снижение сократительной способности мышцы до полного отсутствия. Оптимум – такая частота раздражителя, при которых амплитуда мышечного сокращения максимальна (каждое последующее раздражение наносится в фазу повышенной возбудимости). Пессимум – чрезмерно большая частота раздражений, которая ведет к резкому уменьшению амплитуды мышечного сокращения или к отсутствию реакции (каждое последующее раздражение наносится в фазу пониженной возбудимости). Лабильность. Парабиоз и его фазы. (Н.Е. Введенский). Нервные волокна обладают лабильностью — способностью воспроизводить определенное количество циклов возбуждения в единицу времени в соответствии с ритмом действующих раздражителей. Мерой лабильности является максимальное количество циклов возбуждения, которое способно воспроизвести нервное волокно в единицу времени без трансформации ритма раздражения. Лабильность определяется длительностью пика потенциала действия, т. е. фазой абсолютной рефрактерности. Так как длительность абсолютной рефрактерности у спайкового потенциала нервного волокна самая короткая, то лабильность его самая высокая. Нервное волокно способно воспроизвести до 1000импульсов в секунду. Явление парабиоза открыто русским физиологом Н.Е. Введенским в 1901 г. при изучении возбудимости нервно-мышечного препарата. Состояние парабиоза могут вызвать различные воздействия – сверхчастые, сверхсильные стимулы, яды, лекарства и другие воздействия как в норме, так и при патологии. Н. Е. Введенский обнаружил, что если участок нерва подвергнуть альтерации (т. е. воздействию повреждающего агента), то лабильность такого участка резко снижается. Восстановление исходного состояния нервного волокна после каждого потенциала действия в поврежденном участке происходит медленно. При действии на этот участок частых раздражителей он не в состоянии воспроизвести заданный ритм раздражения, и поэтому проведение импульсов блокируется. Такое состояние пониженной лабильности и было названо Н. Е. Введенским парабиозом. Состояние парабиоза возбудимой ткани возникает под влиянием сильных раздражителей и характеризуется фазными нарушениями проводимости и возбудимости. Выделяют 3 фазы: первичную, фазу наибольшей активности (оптимум) и фазу сниженной активности (пессимум). Третья фаза объединяет 3 последовательно сменяющие друг друга стадии: уравнительную (провизорная, трансформирующая – по Н.Е. Введенскому), парадоксальную и тормозную. Первая фаза (примум) характеризуется снижением возбудимости и повышением лабильности. Во вторую фазу (оптимум) возбудимость достигает максимума, лабильность начинает снижаться. В третью фазу (пессимум) возбудимость и лабильность снижаются параллельно и развивается 3 стадии парабиоза. Первая стадия - уравнительная по И.П. Павлову - характеризуется выравниванием ответов на сильные, частые и умеренные раздражения. В уравнительную фазу происходит уравнивание величины ответной реакции на частые и редкие раздражители. В нормальных условиях функционирования нервного волокна величина ответной реакции иннервируемых им мышечных волокон подчиняется закону силы: на редкие раздражители ответная реакция меньше, а на частые раздражители—больше. При действии парабиотического агента и при редком ритме раздражении (например, 25 Гц) все импульсы возбуждения проводятся через парабиотический участок, так как возбудимость после предыдущего импульса успевает восстановиться. При высоком ритме раздражении (100Гц) последующие импульсы могут поступать в тот момент, когда нервное волокно еще находится в состоянии относительной рефрактерности, вызванной предыдущим потенциалом действия. Поэтому часть импульсов не проводится. Если проводится только каждое четвертое возбуждение (т.е. 25 импульсов из 100), то амплитуда ответной реакции становится такой же, как на редкие раздражители (25Гц)—происходит уравнивание ответной реакции. Вторая стадия – парадоксальная, характеризуется извращенным реагированием – сильные раздражения вызывают меньший ответ, чем умеренные. В эту - парадоксальную фазу происходит дальнейшее снижение лабильности. При этом на редкие и частые раздражители ответная реакция возникает, но на частые раздражители она значительно меньше, т. к. частые раздражители еще больше снижают лабильность, удлиняя фазу абсолютной рефрактерности. Следовательно, наблюдается парадокс— на редкие раздражители ответная реакция больше, чем на частые. Третья стадия – тормозная. Фазулабильность снижается до такой степени, что и редкие, и частые раздражители не вызывают ответной реакции. При этом мембрана нервного волокна деполяризована и не переходит в стадию реполяризации, т. е. не восстанавливается ее исходное состояние. Ни сильные, ни умеренные раздражения не вызывают видимой реакции, в ткани развивается торможение. Имеет место четвертая — ультрапарадоксальная, при которой положительные раздражители вызывают отрицательный эффект, а отрицательные — положительный. Парабиоз— явление обратимое. Если парабиотическое вещество действует недолго, то после прекращения его действия нерв выходит из состояния парабиоза через те же фазы, но в обратной последовательности. Однако, при действии сильных раздражителей за тормозной стадией может наступить полная потеря возбудимости и проводимости, а в дальнейшем – гибель ткани. Работы Н.Е. Введенского по парабиозу сыграли важную роль в развитии нейрофизиологии и клинической медицины, показав единство процессов возбуждения, торможения и покоя, изменили господствовавший в физиологии закон силовых отношений, согласно которому реакция тем больше, чем сильнее действующий раздражитель. Явление парабиоза лежит в основе медикаментозного локального обезболивания. Влияние анестезирующих веществ вязано с понижением лабильности и нарушением механизма проведения возбуждения по нервным волокнам. Типы мышечных волокон. Нейромоторные единицы и их особенности в разных мышцах. Классификации мышечных волокон В настоящее время общепринято считать, что у человека скелетные мышцы состоят из волокон различных типов. Существуют различные классификации типов мышечных волокон. Различают волокна: красные и белые, медленные и быстрые, тонические и фазические. В середине ХХ века для разделения мышечных волокон на разные типы использовались гистологические методы (А.В. Самсонова с соавт., 2012). Из скелетных мышц посредством биопсии извлекался кусочек мышечной ткани, быстро замораживался и разрезался на тонкие слои. Затем производилось исследование мышечной ткани под микроскопом. Первоначально критерием разделения мышечных волокон на медленные и быстрые являлось количество и расположение митохондрий. Затем предпочтение стали отдавать такому показателю как толщина Z-дисков. Было найдено, что у медленных волокон Z-диски существенно толще, чем у быстрых. В качестве еще одного критерия разделения мышечных волокон на типы использовалась толщина М-диска. При продольных срезах расслабленной скелетной мышцы видно, что медленные мышечные волокна содержат пять М-линий, имеющих одинаковую плотность. Промежуточные мышечные волокна – три линии средней плотности, ясно видимые и две линии, имеющие небольшую плотность. В быстрых мышечных волокнах имеются три линии средней плотности и две внешние, едва различимые. В настоящее время чаще всего используется классификация M.Brook, K.Kaiser (1970), которая основывается на гистохимических методах. Известно, что миофибриллы состоят из саркомеров, а те, в свою очередь из толстых и тонких филаментов. Основу толстых филаментов составляет белок миозин, а основу тонких – белок актин. Гистохимические методы основаны на определении активности фермента АТФ-азы миозина. Этот фермент расположен на головках молекул миозина. Фермент АТФ-аза осуществляет высвобождение энергии, необходимой для осуществления сокращения мышечного волокна. Степень активности АТФ-азы варьирует в широких пределах. Установлено, что степень активности АТФ-азы миозина связана с типом миозина, содержащемся в мышечном волокне. В медленных мышечных волокнах активность АТФ-азы низкая, а в быстрых – высокая. Именно высокая активность АТФ-азы миозина способствует высокой скорости сокращения мышечных волокон. На основе классификации по активности АТФ-азы миозина различают мышечные волокна типа I, типа IIA и типа IIB. Характеристики различных типов (видов) мышечных волокон Медленные и быстрые мышечные волокна различаются метаболизмом, что проявляется в активности ферментов и количестве митохондрий. Медленные мышечные волокна окружены большим числом крупных митохондрий с набором ферментов, катализирующих распад углеводов и жирных кислот. Поскольку этот процесс требует притока большого количества кислорода, вполне естественно, что сеть капилляров, окружающая медленные мышечные волокна более развита и снабжение кислородом, доставленным с током крови, в этих волокнах происходит более интенсивно. В этих волокнах крайне ограничен запас углеводов в виде гликогена и низка активность ферментов гликолиза (М.И. Калинский, В.А. Рогозкин, 1989). Быстрые волокна типа IIA и IIB характеризуются высокой активностью АТФ-азы миозина, поэтому скорость их сокращения практически в два раза выше, чем у медленных. С высокой скоростью сокращения связан хорошо развитый саркоплазматический ретикулум, который характерен для быстрых мышечных волокон, так как он содержит ионы кальция, необходимые для сокращения мышечного волокна. Волокна типа IIA имеют набор ферментов для полного окисления углеводов и жирных кислот, такой же, как и в медленных волокнах и к тому же они располагают ферментами гликолиза, то есть способностью расщеплять углеводы до молочной кислоты. Быстрые мышечные волокна типа IIB способны к коротким периодам сократительной активности. Они имеют набор ферментов гликолиза с высокой активностью и небольшое количество митохондрий с окислительными ферментами. Быстрые мышечные волокна типа IIA и IIB имеют большие запасы гликогена, который сразу используется в качестве источника энергии при сокращении скелетной мышцы (табл.1). Таблица 1 Характеристики мышечных волокон различных типов
Функции мышечных волокон Основная функция волокон типа I – выполнение длительной работы низкой интенсивности. Они активны также при поддержании позы. Поэтому антигравитационные мышцы в основном состоят из медленных волокон типа I. Основная функция мышечных волокон типа II – выполнение быстрых и сильных сокращений. Расположение мышечных волокон различных типов в скелетных мышцах Мышечные волокна объединены в пучки. Их покрывает перимизий. Пучок содержит мышечные волокна различных типов. В пучке мышечные волокна расположены в виде мозаики. Однако доказано, что внутри мышцы больше мышечных волокон типа I, а снаружи – мышечных волокон типа II. Понятие о нейромоторных единицах. Нейромоторная единица - совокупность нейрона и группы мышечных волокон, иннервируемых аксоном этого нейрона. В состав нейромоторной единицы входят: нервная клетка - в основном мотонейроны, тела которых лежат в передних рогах спинного мозга; аксон мотонейрона - миелиновые волокна; группа мышечных волокон - в зависимости от вида деятельности количества волокон различно. Если тонкая работа 2-4, если грубая - до нескольких тысяч. Виды нейромоторных единиц. По характеру возбуждения, возникающего в мышечных волокнах все нейромоторные единицы, делятся на 2 группы. Фазные нейромоторные единицы - образуются альфа-мотонейронами. Это самые крупные мотонейроны (диаметр 10-20 мкм). Скорость проведения возбуждения по отростку этого аксона - 120 м/с. Аксон обеспечивает одиночную иннервацию, образуя на каждом волокне 1-2 синапса. В каждом синапсе при возбуждении возникает достаточный потенциал концевой пластинки, который обеспечивает возникновение потенциала действия в мышце (импульсное возбуждение). Лучше работают под действием одиночного возбуждения (импульса), при котором возникает распространяющееся возбуждение. Среди фазных нейромоторных единиц выделяют быстрые и медленные. Быстрые - продолжительность потенциала действия в 2 раза меньше, чем в медленных. Волна сокращения в 5 раз меньше, чем в медленных. Скорость распространения возбуждения в быстрых нейромоторных единицах в 2 раза больше, чем в медленных. Таким образом, быстрые фазные нейромоторные единицы обеспечивают динамическую работу, когда быстрое сокращение сменяется быстрым расслаблением. Для этого вида нейромоторных единиц характерно анаэробное образование энергии. Эти мышечные волокна практически не содержат миоглобина - светлые, белые мышцы; Медленные - обеспечивают в основном статическую работу, медленное, длительное сокращение мышц. Основной поставщик энергии окислительно-восстановительные процессы. Содержит миоглобин, который депонирует кислород. По цвету темные, красные мышцы. Тонические нейромоторные единицы - образуются гамма-мотонейронами передних рогов спинного мозга (самые мелкие, диаметр 4-6 мкм). Скорость проведения импульса по отросткам этих нейронов - 30 м/с, в синапсах возникает местный потенциал концевой пластинки (местное возбуждение). Аксоны тонических нейронов обеспечивают множественную иннервацию, т. е. на каждом мышечном волокне образуются несколько десятков синапсов и за счет суммации местных потенциалов и возникает потенциал действия. Возбуждение тонической нейромоторной единицы происходит под влиянием серии импульсов (частота около 10 Гц). Тонические нейромоторные единицы обеспечивают медленное сокращение мышц, участвуют в возникновении тонуса. Скелетные мышцы образуются различными мышечными волокнами, которые входят в состав фазных и тонических нейромоторных единиц. Попеременное включение нейромоторных единиц обеспечивает изменение функционального состояния мышцы. В состоянии покоя работают тонические нейромоторные единицы, в состоянии активности - фазные нейромоторные единицы. Кроме двигательной иннервации присутствует вегетативная. Все скелетные мышцы получают импульсы из симпатической нервной системы, которая регулирует обменные процессы. Тонус мышц. Механизм возникновения мышечного тонуса. Тонус - умеренное напряжение мышц, когда они находятся в состоянии относительного покоя. Тонус поддерживается за счет нервных импульсов, поступающих из центральной нервной системы даже в состоянии покоя. Источники импульсов - мотонейроны (альфа и гамма) передних рогов спинного мозга. Они должны сами находиться в состоянии тонуса. Причины тонуса нейронов - рефлекторное происхождение тонуса мышц - мотонейроны получают импульсы от рецепторов скелетных мышц. Доказательство: исчезновение тонуса скелетных мышц при перерезке задних корешков спинного мозга; действие гуморальных факторов - активность мотонейронов поддерживается за счет действия метаболитов (например, СО2, лактат - накапливается в спинном мозге, возбуждая нейроны); влияние вышележащих отделов центральной нервной системы - они поддерживают тонус мышц и регулируют его уровень и распределение. Доказательство: удаление головного мозга у лягушки. Сила и работа мышц. Закон средних нагрузок. Величина сокращения (степень укорочения) мышцы при данной силе раздражения (т. е. при данном числе активированных волокон) зависит как от ее морфологических свойств, так и от физиологического состояния. Длинные мышцы сокращаются на большую величину, чем короткие. Умеренное растяжение мышцы увеличивает ее сократительный эффект; при сильном растяжении сокращение мышцы ослабляется. Если в результате длительной работы развивается утомление мышцы, то величина ее сокращения снижается. Для измерения силы мышцы определяют либо тот максимальный груз, который она в состоянии поднять, либо максимальное напряжение, которое она может развить в условиях изометрического сокращения. Эта сила может быть очень велика. Установлено, что собака мышцами челюсти может поднять груз, превышающий массу ее тела в 8,3 раза. Одиночное мышечное волокно способно развить напряжение 100—200 мг. Учитывая, что общее количество мышечных волокон в теле человека приблизительно 15—30 млн., они могли бы развить напряжение в 20—30 т, если бы все одновременно тянули в одну сторону. Сила мышцы при прочих равных условиях зависит от ее поперечного сечения. Чем больше физиологическое поперечное сечение мышцы, т. е. сумма поперечных сечений всех ее волокон, тем больше тот груз, который она в состоянии поднять. Физиологическое поперечное сечение совпадает с геометрическим только в мышцах с продольно расположенными волокнами; у мышц с косым расположением волокон сумма поперечных сечений может значительно превышать геометрическое поперечное сечение самой мышцы. По этой причине сила мышцы с косо расположенными волокнами значительно больше, чем сила мышцы той же толщины, но при продольном расположении волокон. Чтобы иметь возможность сравнивать силу разных мышц, максимальный груз, который в состоянии поднять мышца, делят на площадь ее физиологического поперечного сечения. Таким образом вычисляют абсолютную мышечную силу. Абсолютная сила, выраженная в килограммах на I см2, икроножной мышцы человека равна 5,9, сгибателя плеча — 8,1, жевательной мышцы — 10, двуглавой мышцы плеча — 11,4, трехглавой мышцы плеча — 16,8, гладких мышц— 1. Большинство мышц млекопитающих и человека перистогофизиологическое сечение, а потому обладает большой силой. Работа мышцы измеряется произведением поднятого груза на величину укорочения мышцы. Между грузом, который поднимает мышца, и выполняемой ею работой существует следующая зависимость. Внешняя работа мышцы равна нулю, если мышца сокращается без нагрузки. По мере увеличения груза работа сначала увеличивается, а затем постепенно уменьшается. При очень большом грузе, который мышца неспособна поднять, работа становится равной нулю. Наибольшую работу мышца совершает при некоторых средних нагрузках, в данном случае при 200—250 г. Мощность мышцы, измеряемая величиной работы в единицу времени, также достигает максимальной величины при средних нагрузках. Поэтому зависимость работы и мощности от нагрузки получила название правила средних нагрузок. Для того, чтобы понять, почему мышца выполняет максимальную работу при средних нагрузках, необходимо рассмотреть состояние саркомера при разной степени растяжения мышцы. Если покоящуюся мышцу постепенно растягивать за сухожильные концы, то в силу эластических свойств напряжение мышцы будет возрастать. Нанесение электрических раздражений на мышцу вызывает прирост напряжения, величина которого с увеличением исходной длины мышечных волокон вначале возрастает, а затем уменьшается. При некоторой средней длине саркомеров сила, развиваемая мышечным волокном, максимальна. При перерастяжении волокон эта сила уменьшается. Таким образом, мышца развивает максимальную работу при условии наибольшего перекрытия миозиновых и актиновых нитей. Функциональная характеристика гладких мышц. Гладкие мышцы находятся в стенках внутренних органов и кровеносных сосудов. Регуляция их тонуса и сократительной активности осуществляется эффективными волокнами симпатической и парасимпатической НС, а также местными гуморальными и физическими воздействиями. Сократительный аппарат гладких мышц, как и скелетных, состоит из толстых миозиновых и тонких актиновых нитей. Вследствии их нерегулярного распределения клетки гладких мышц не имеют характерной для скелетной и сердечной мышцы поперечной исчерченности. Гладкомышечные клетки имеют веретенообразную форму, l = 500-400 мкм. и толщину 2-10 мкм. Отделены друг от друга узкими щелями (60-150 нм). Возбуждение электротонически распространяется по мышце от клетки к клетке через особые контакты (нексусы) между плазматическими мембранами соседних клеток. Гладкая мышца с морфологической точки зрения является не истинным, а функциональным синцитием. Клетки в гладкой мышце расположены хаотично, неравномерно, иногда они собраны в пучки или тяжы. Их окружает соединительная ткань. Волокна гладких мышц сокращаются в результате относительного скольжения миозиновых и атктиновых нитей, но скорость их сокращения и скорость расщепления АТФ в 100-1000 раз меньше, чем в скелетных мышцах. Гладкие мышцы хорошо приспособлены к длительному тоническому сокращению без развития утомления. При этом их энерготраты крайне невелики. По функциональным особенностям гладкие мышцы подразделяются на мышцы, обладающие и не обладающие спонтанной активностью. Гладкие мышцы, обладающие спонтанной активностью способны сокращаться и при отсутствии прямых, возбуждающих нервных и гуморальных воздействий (например, ритмические сокращения гладких мышц кишечника). ПП таких клеток постоянно спонтанно колеблются в пределах 30-70 мВ («дрейф»). При снижении потенциала покоя до критического уровня возникает потенциал действия, вызывающий сокращение мышечного волокна. Длительность ПД гладкомышечных волокон намного больше, чем у волокон поперечно-полосатой мускулатуры (несколько сек.) Продолжительность самого сокращения также превышает несколько секунд. Особенно медленно протекает расслабление. Возбуждение распространяется со скоростью 2-10 см/сек, т.е. меньшей, чем по скелетной мышце. Спонтанная активность гладкомышечных клеток связана с их растяжением, вызывающим деполяризацию мембраны мышечных клеток серии импульсов с последующим сокращением. Гладкие мышцы, не обладающие спонтанной активностью, сокращаются под влиянием импульсов вегетативной НС. Так, в отличие от мышц кишечника, мышечные клетки артерий, семенных протоков и радужки обладают слабой спонтанной активностью, или вообще не проявляют ее. Отдельные нервные импульсы не способны вызвать пороговую деполяризацию таких клеток и их сокращения. ПД волокна с последующим сокращением возникает лишь при поступлении к нему серии импульсов с частотой 1 импульс в секунду и выше. Возбуждение передается от одной клетки к последующим через плотные контакты их мембран. Виртуальные лабораторные работы: Раздел «Физиология мышечной системы» Простое сокращение скелетной мышцы Роль нейромышечного синапса в возникновении утомления Сокращение скелетных мышц в результате действия нескольких стимулов. Лабораторные работы: №29 Определение двигательной хронаксии человека (см. прикрепленный файл) №31 Электромиограмма* См. Лабораторный практику по физиологии человека. Киселев В.Д., Томилова И.Н., Плешкова Н.В. – 2009. – 187с. |