Возбудимость, проводимость, автоматизм
 Скачать 3.49 Mb. Скачать 3.49 Mb.
|

Электрофизиология нервного стволаНервный ствол, или нерв, представляет собой пучок множества нервных волокон, покрытых у позвоночных общими эпителиальной и соединительнотканной оболочками. Обычно нерв включает в себя волокна различного типа и разного диаметра. Например, седалищный нерв лягушки содержит двигательные волокна (аксоны мотонейронов), чувствительные (аксоны чувствительных клеток) и автономные (аксоны нейронов симпатических ганглиев). Моторные и наиболее толстые чувствительные волокна этого нерва миелинизированы, симпатические — не имеют миелиновой оболочки, покрыты одним слоем шванновских клеток. Все эти волокна в покое обладают поляризованной поверхностной мембраной. Немецкий физиолог Эмиль Дю Буа-Реймон показал, что если нерв перерезать и соединить его поврежденную и продольную поверхность с регистрирующим устройством, то отводится потенциал повреждения, или потенциал покоя (ПП). [++512+с374]. Потенциал нормальной точки нерва относительно поврежденной имеет обычно небольшую величину — 20—30 мВ, он меньше ПП волокон нерва, что объясняют шунтирующим действием межволоконных щелевых пространств. Если в сегменте нерва, расположенном между электродами, заменить (перфузией) межклеточную жидкость на раствор сахарозы (сахарозный мостик), то ПП нерва «поднимается» до —70 мВ. Его величина, по существу, отражает величину МПП наиболее толстых волокон, генерирующих основную часть внешнего тока.[++421+c62]. Вспомним лекцию по законам раздражения. Нервный ствол является составной возбудимой структурой отвечающей на раздражение между минимальным и максимальным порогом по закону «силы», в то время как составляющие его элементарные возбудимые структуры (нервные волокна) отвечают на раздражение по закону «всё или ничего» (рис. 210041815).  Рис. 210041815. Закон силы раздражения в приложении к составной возбудимой структуре (нерву, мышце).. Почему это происходит? Нервные волокна имеют разный порог раздражения. Когда фиксируется минимальный порог раздражения? Когда возбудились волокна имеющие наименьшие пороги раздражения. При увеличении силы стимула всё большее количество волокон вовлекается в реакцию возбуждения. При достижении максимального порога преодолеваются пороги раздражения всех волокон. При раздражении нервного стола естественно должно быть воздействие электрическим током через наружные (внеклеточные) электроды. При этом нужно учитывать ветвление тока в нерве, шунтирование тока в межклеточных щелях (рис. 210041846).  Рис. 210041846. Ветвление тока в ткани при раздражении через наружные (внеклеточные) электроды (схема). Волокна обведены толстой линией, между ними – межклеточные щели. Объяснение в тексте. Классификация нервных волокон по Эрлангеру-ГассеруВ 1939 г. американские физиологи Джозеф Эрлангер и Герберт С.Гассер зарегистрировали токи действия от целого нервного ствола седалищного нерва лягушки на разных расстояниях от стимулирующего электрода (рис. 210041905). Было установлено, что регистрируемый суммарный потенциал имеет ряд пиков, которые были обозначены буквами латинского алфавита A, B, C (рис. , I). Пик A имел дополнительные пики, помеченные греческими буквами α, β, γ, δ (рис. , II). В 1944 г. работа Дж.Эрлангера и Г.С.Гассера была оценена присуждением Нобелевской премии.  Нервы у позвоночных состоят из трех основных групп волокон (А, В и С), различающихся по степени миелизации, диаметру волокна, длительности пика ПД (скорости развития ПД), электровозбудимости, его компенсации и скорости проведения (все эти показатели в ряду А — В — С падают). Группа А включает наиболее толстые хорошо миелинизированные моторные и чувствительные волокна; группа В — слабомиелинизированные, преганглионарные волокна автономной нервной системы; группа С - немиелинизированные, постганглионарные (симпатические) волокна. В группе А в ряду , , , названные показатели тоже падают. Соотношения свойств этих групп волокон демонстрируются в табл. . Необходимо заметить, что указанные соотношения порогов электрического раздражения групп волокон не отражают точного соотношения электровозбудимости их мембран. Относительно высокие пороги тонких волокон при их раздражении в нервном стволе определяются в основном тем обстоятельством, что тонкие волокна по сравнению с толстыми обладают более высоким входным сопротивлением. В них входит такая малая часть раздражающего тока, что при пороговой силе для А-волокон она совершенно недостаточна для создания на мембране более тонких волокон сколько-нибудь существенной деполяризации. По этой же причине (высокое RI) отводимые от ствола (внеклеточно) ПД тонких волокон предстают значительно меньшими, чем ПД толстых волокон. 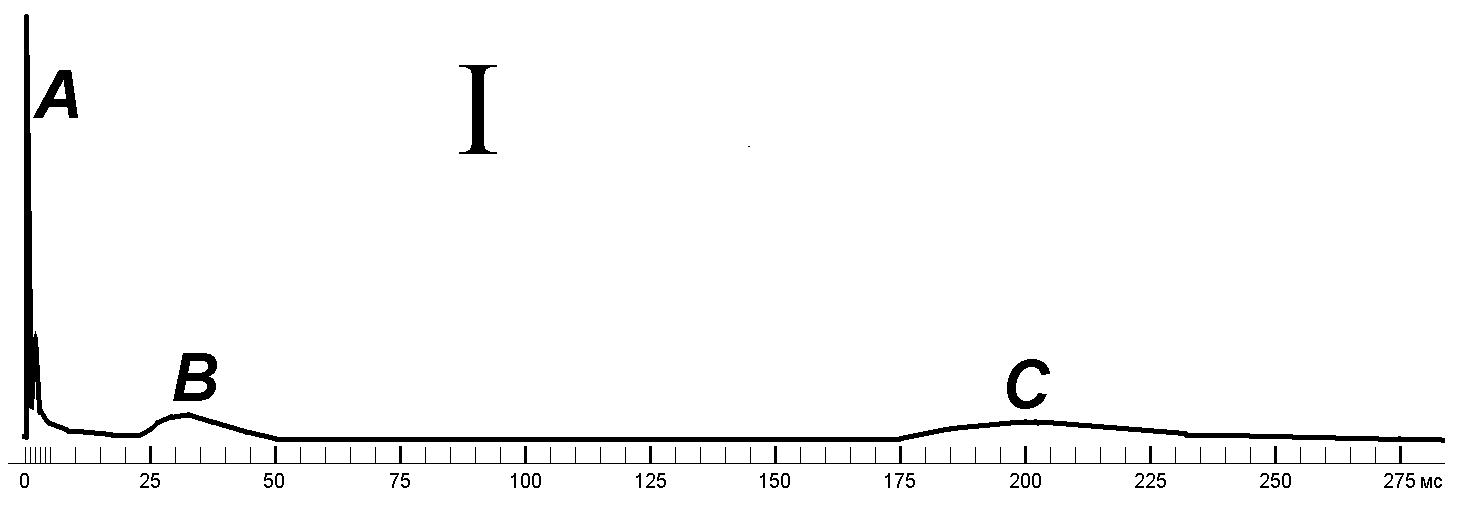 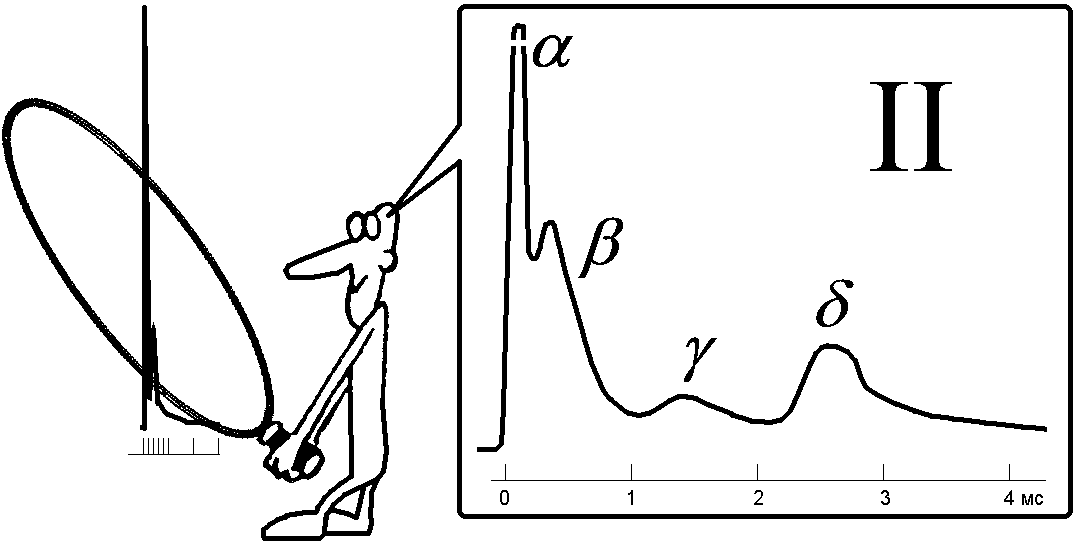 Рис. . Составные части потенциала действия смешанного нерва. I – при относительно медленной скорости записи. II – при относительно высокой скорости записи. Объяснение в тексте. По оси абсцисс – время, по оси ординат амплитуда составного потенциала в мВ. Суммарная электрическая активность нерва создается его волокнами, каждое из которых генерирует свой стандартный по амплитуде и временным параметрам ПД, распространяющийся в обе стороны от точки, к которой приложено раздражение. Суммарный электрический сигнал нерва зависит от числа активных волокон, синхронности их активности, способа отведения и других обстоятельств. Рассмотрим случай так называемого однофазного отведения, при котором один отводящий электрод (активный) расположен на нормальном участке ствола, а другой (индифферентный) — на поврежденном, где волокна деполяризованы полностью (рис.1.19). Пусть применяют искусственное раздражение и раздражающий электрод (катод) находится достаточно близко (l= 3 мм) от активного отводящего электрода, а нерв помещен в непроводящую среду (масло или воздух). Здесь сильное одиночное раздражение приводит к синхронному возбуждению всех волокон, при этом активный отводящий электрод регистрирует суммарный ПД нерва, по форме приближающийся к ПД отдельного A(альфа)‑волокна, но немного более затянутый во времени. Этот ПД нерва, однако, не подчиняется правилу «все или ничего». При пороговом раздражении он ничтожно мал, с увеличением силы стимулов постепенно растет, достигая максимума, равного при обычных условиях 5—10 мВ, а в условиях сахарозного мостика — 50—100 мВ. При дальнейшем увеличении силы стимула этот ПД несколько удлиняется во времени. Все изменения амплитуды и длительности пика ПД нерва при усилении стимула определяются ростом числа активных волокон, подключением к низкопороговым и быстрым А(альфа)-волокнам более высокопороговых медленных бета-, гамма-, дельта-волокон группы А, затем В- и, наконец, С-группы.
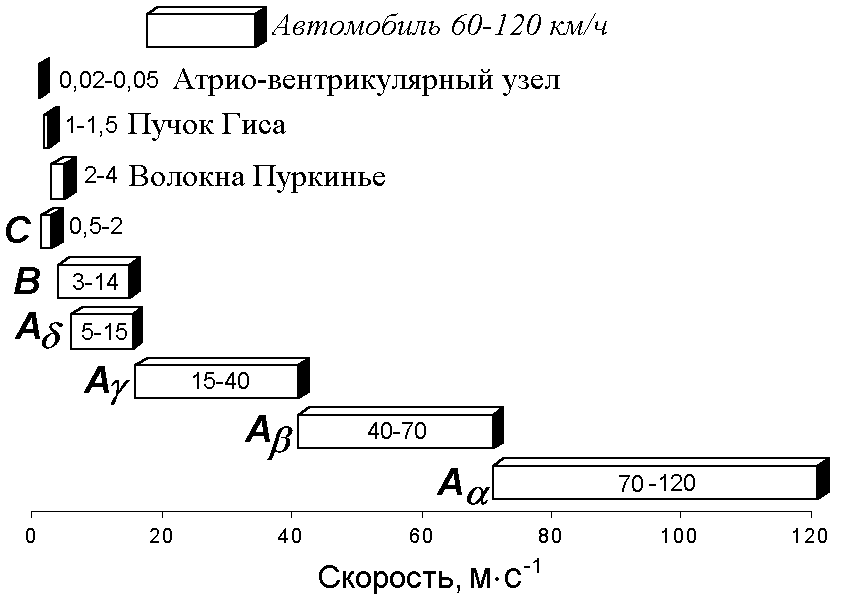 Таблица . Классификация нервных волокон по Дж.Эрлангеру и Х.Гассеру
1Приблизительно ту же величину имеют и абсолютные рефрактерные фазы | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||