Конспекты по физиологии. Конспект по физиологии 8-10. Тема Физиология мышечного сокращения
 Скачать 113.38 Kb. Скачать 113.38 Kb.
|

|
Тема 8. Физиология мышечного сокращения Структурной единицей мышечного волокна являются миофибриллы. Они разделены на чередующиеся участки (диски), которые обладают различными оптическими свойствами. Диски, обладающие двойным лучепреломлением, получили название анизотропные (А) диски. Диски, которые не обладают двойным лучепреломлением, названы изотропные (I) диски. Анизотропные диски в обыкновенном свете выглядят темными и состоят из двух темных полосок, разделенных светлой "H" полоской. Изотропные диски в обыкновенном свете выглядят светлыми и в середине имеют темную "Z" полоску. Z полоска – эта тонкая мембрана, которая является продолжением поверхностной мембраны вглубь мышечного волокна. Она выполняет опорную функцию, поскольку через ее поры проходят протофибриллы. В зоне Z мембраны также находятся триады или Т-системы триады представляют выпячивания плазматической мембраны с образованием поперечных трубочек в виде ярусов и цисцерн. Они предсталяют саркоплазматический ретикулум, который содержит высокую концентрацию ионов Ca. При возбуждении Z мембраны кальций по концентрационному градиенту выходит из саркоплазматического ретикулума в протофибриллярное пространство, вызывая процесс сокращения Активная реабсорбция ионов Са в саркоплазматический ретикулум за счет работы Са-насоса, приводит к расслаблению мышечного волокна. Структурной единицей миофибриллы являются протофибриллы Протофибриллы включают белковые нити актина и миозина, а также белки тропонин и тропомиозин. Нити миозина – это толстые и короткие нити, которые входят только в состав анизотропного диска. Нити актина – это тонкие и длинные нити, входящие в состав как изотропного, так и анизотропного дисков. Они вставлены между нитями миозина. От них свободна только H-полоска анизотропного диска. Процесс сокращения происходит в результате скольжения нитей актина относительно нитей миозина, который запускается накоплением Са, при этом образуются актино-миозиновые комплексы (мостики) и нити актина вдвигаются в промежутки между нитями миозина. Нити актина сближаются друг с другом. Ширина А-диска (1,6 мкм) всегда остаётся постоянной, тогда как I-диски и H-полоски при сокращении сужаются Механизм мышечного сокращения и расслабления. 1. Раздражение. 2. Возникновение потенциала действие. 3. Проведение возбуждения вдоль клеточной мембраны до Z мембраны, а далее вглубь волокна по трубочкам саркоплазматического ретикулума. 4. Освобождение Са из триад. 5. Диффузия Са к протофибриллам. 6. Взаимодействие Са с тропонином. 7. Конформационное изменение комплекса тропомиозин-тропонин. 8. Освобождение активных центров актина. 9. Присоединение актина к миозину. 10. В присутствии белка актомиозина распад АТФ с освобождением энергии. 11. Скольжение нитей актина относительно миозина. 12. Укорочение миофибриллы. 13. Активация кальциевого насоса. 14. Ресинтез АТФ. 15. Понижение концентрации свободных ионов Са в саркоплазме. 16. Разрушение актин-миозиновых комплексов. 17. Обратное скольжение нитей актина относительно миозина. 18. Увеличение (восстановление) миофибриллы. Мышечные веретена принадлежат к группе наиболее изученных рецепторов. Под световым микроскопом мышечное веретено представляет собой вытянутую структуру, расширенную в середине за счет капсулы и напоминающее по форме веретено, что и обусловило его название. Мышечные веретена у всех видов животных имеют примерно одинаковое строение (рис. 15.2). Вспомогательные структуры мышечных волокон представлены специализированными поперечнополосатыми мышечными волокнами, названными интрафузалъными (в отличие от обычных, экстрафузальных волокон), а также капсулой рецептора, за- 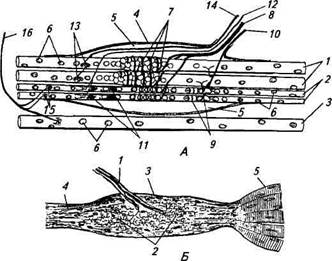 Рис. 15.2. Строение мышечного веретена (А) и сухожильного ре цептора Гольджи (Б): А. 1 — интрафузальное мышечное волокно с ядерной сумкой; 2 — интра-фузальное мышечное волокно с ядерной цепочкой; 3 — экстрафузальное мышечное волокно; 4— капсула; 5 — внутрикапсулярное пространство; 6— ядра мышечных волокон; 7—первичные афферентные окончания; 8— афферентное нервное волокно первого типа; 9— вторичные нервные окончания; 10— афферентное нервное волокно второго типа; 11 — у-кус-тиковидные эфферентные нервные окончания; 12— у2-двигательное нервное волокно; 13 — у-пластинчатые эфферентные нервные окончания; 14 — у [-двигательное нервное волокно; 15 — р-пластинчатые эфферентные нервные окончания; 16— р-двигательное нервное волокно; Б. 1 — афферентное нервное волокно; 2— афферентные нервные окончания; 3— капсула; 4— сухожилия; 5— мышечные волокна полненной жидкостью. В полярных областях интрафузальных волокон хорошо видны ядра, рассеянные по периферии волокна, как и в экстрафузальных. Ближе к центральной части, где волокно покрыто капсулой, появляются дополнительные ядра. Это и послужило основанием называть центральную зону волокна областью ядерной сумки, а сами интрафузальные волокна называют волокнами с ядерной сумкой; диаметр этих волокон составляет 20...25мкм. У более тонких волокон другого типа (диаметр около 10 мкм) ядра располагаются в ряд, образуя цепочку. Такие волокна получили название интрафузальных волокон с ядерной цепочкой. Интрафузальные волокна покрыты соединительнотканной капсулой. В центральной области веретена, где капсула отходит от интрафузальных волокон, имеется перикапсулярное пространство, заполненное жидкостью. К мышечному веретену подходит большое количество нервных волокон, разветвляющихся в разных его частях. той капсулой. В мышечных веретенах было обнаружено штышьт типов афферентных и эфферентных двигательных окончании Последние согласно общей схеме строения анализатором можно отнести к звену обратной связи. Афферентные окончания представлены двумя типами. 11 с р вый тип (первичные окончания) образуется разветвлени ями довольно толстых миелинизированных волокон, достигающих в диаметре около 20 мкм. Нервное волокно проникает под капсулу и, многократно разветвляясь, постепенно теряет миелиновую оболочку. Первичные окончания локализуются в середине центрального участка интрафузальных волокон и имеют вид спиралей и колец, обвивающих волокна. Второй тип (вторичные окончания) образуется более тонкими миелинизи-рованными волокнами (диаметр 4... 12 мкм) и располагается ближе к периферии интрафузального волокна (см. рис. 15.2). Диаметр обоих типов окончаний в поперечном сечении составляет 0,5...3 мкм. Характерная их особенность — это большое число митохондрий во внутриклеточном пространстве. Мышечные веретена иннервируют также и несколько типов двигательных нервных волокон, получивших название фузимоторных (у-волокон). Фузимоторные синаптические окончания, образованные этими волокнами, подразделяются на у-кустиковид-ные и у-пластинчатые. Первые локализуются ближе к центру веретена, а у-пластинчатые — дистальнее. Кроме двигательных у-волокон мышечные веретена дополнительно иннервируют двигательные Р-волокна, которые одновременно образуют синаптические окончания на экстрафузальных мышечных волокнах, р-волокна на интрафузальных волокнах формируют окончания пластинчатого типа, располагающиеся в самых ди-стальных областях интрафузальных волокон. В исследованиях реакции интрафузальных мышечных волокон на раздражение эфферентных волокон было обнаружено, что способностью к сокращению обладают только дистальные участки, а центральная часть несократима. При регистрации электрической активности в афферентных нервных волокнах, идущих от мышечных веретен, в состоянии покоя были обнаружены нерегулярные низкочастотные потенциалы действия, получивших название спонтанных. Принимая во внимание механизмы генерации потенциалов действия в нервных клетках и волокнах, можно полагать, что мембрана рецеп-торной области исходно деполяризована и значение мембранного потенциала находится в области порога возникновения потенциалов действия. Таким образом, даже самое маленькое увеличение энергии раздражающего стимула (в данном случае механического) будет повышать частоту импульсации рецептора и его чувствительность очень высока. Растяжение мышечного веретена приводит к возникновению электрической активности — рецепторного потенциала и потенциалов действия. При легком ступенчатообразном растяжении мышечного веретена в первичных и во вторичных афферентных окончаниях регистрируется длительная серия потенциалов действия. Частота разрядов зависит от степени растяжения мышцы. На рисунке видно, что потенциалы действия находятся на своеобразном деполяризационном пьедестале, представляющем собой рецепторный потенциал, т. е. изменение мембранного потенциала афферентных окончаний. Более сильное растяжение приводит к более значительной деполяризации и увеличению частоты афферентных потенциалов действия. Частота потенциалов действия мышечного веретена на фоне неизменной амплитуды растяжения изменяется со временем. Начало растяжения сопровождается повышением частоты импульсов с последующим быстрым спадом до некоторого уровня (динамическая часть ответа). Далее частота продолжает уже медленнее снижаться до значения, определяемого величиной растяжения (статическая часть ответа). Процесс снижения частоты потенциалов действия получил название адаптации. Таким образом, в ответе мышечного веретена четко выражены динамическая и статическая фазы, во время которых соответственно происходит быстрая и медленная адаптация Синапс (греч. synapsis — соединение) — это специализированная структура, обеспечивающая передачу сигнала от клетки к клетке. Посредством синапса реализуется действие многих фармакологических препаратов. Структурно-функциональная организация. Каждый синапс имеет пре- и постсинаптическую мембраны и синаптическую щель . 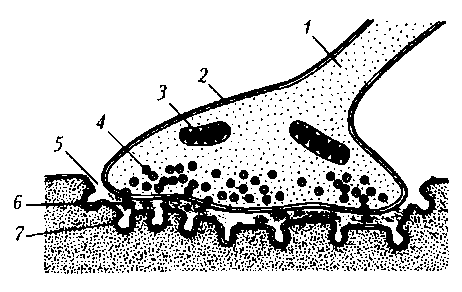 Рис. 17. Нервно-мышечный синапс скелетной мышцы: 1 – ветвь аксона; 2 – пресинаптическое окончание аксона; 3 – митохондрия; 4 – синаптические пузырьки, содержащие ацетилхолин; 5 – синаптическая щель; 6 – молекулы медиатора в синаптической щели; 7 – постсинаптическая мембрана мышечного волокна с N-холинорецепторами Пресинаптическая мембрана нервно-мышечного синапса представляет собой часть мембраны пресинаптического окончания аксона мотонейрона. Через нее осуществляется выброс (экзоцитоз) медиатора (лат. mediator — посредник) в синаптическую щель. В нервно-мышечном синапсе медиатором является ацетилхолин. Медиатор пресинаптического окончания содержится в синаптических пузырьках (везикулах), диаметр которых составляет около 40 нм. Они образуются в комплексе Гольджи, с помощью быстрого аксонного транспорта доставляются в пресинаптическое окончание, где заполняются медиатором и АТФ. В пресинаптическом окончании содержится несколько тысяч везикул, в каждой из которых имеется от 1 тыс. до 10 тыс. молекул химического вещества. Постсинаптическая мембрана (концевая пластинка в нервно-мышечном синапсе) — это часть клеточной мембраны иннервируемой мышечной клетки, содержащая рецепторы, способные связывать молекулы ацетилхолина. Особенность этой мембраны: множества мелких складок, увеличивающих ее площадь и количество рецепторов на ней до 10—20 млн в одном синапсе. Синаптическая щель в нервно-мышечном синапсе имеет ширину в среднем 50 нм. Она содержит межклеточную жидкость, ацетилхолинэстеразу и мукополисахаридное плотное вещество в виде полосок, мостиков, в совокупности образующих базальную мембрану, соединяющую пре- и постсинаптическую мембраны. Хронаксиметрия — это метод, определяющий величину хронаксии, т. е. наименьшего времени, в течение которого раздражитель удвоенной пороговой силы вызовет процесс возбуждения. Для измерения хронаксии пользуются специальным прибором — хронаксиметром, состоящим из источника постоянного тока, набора сопротивлений и приспособлений для дозировки времени действия тока, подающегося на объект. В зависимости от устройства приспособления для дозировки времени действия тока различают маятниковые, конденсаторные и электронные хронаксиметры. Маятниковые хронаксиметры очень громоздки и в клинике мало пригодны. В конденсаторных хронаксиметрах время действия тока регулируется разрядом конденсатора и зависит от его емкости. Раздражение, вызываемое таким хронаксиметром, почти нечувствительно. В электронных хронаксиметрах дозировка длительности действия тока производится при помощи радиоламп. В медицине употребляются приборы, дающие градацию напряжения от 1—2 в до 300 В, а длительность действия — от 0,01 мсек до 40 мсек. Хронаксиметрия применяется для определения дегенерации нерва при травмах различных нервных центров. Исследования хронаксии помогают установить сдвиги возбудимости при воздействии различных факторов: работы, тепла, холода, атмосферного давления и т. д. Хронаксиметрия помогает поставить ранний диагноз или уточнить его, обосновать прогноз, выявить эффективность лечебных мероприятий. Хронаксиметрия имеет большое практическое значение, расширяя возможности электродиагностики Электромиография – метод регистрации электрической активности мышц, возникающей в результате их возбуждения. С помощью ЭМГ изучаются двигательные функции человека и животных В космическом полёте у космонавтов периодически измеряется сила мышц, выносливость и утомляемость с помощью электродинамометра. Сила измеряется как наибольшее усилие, развиваемое мышцей. Для оценки выносливости регистрируется время поддержания половины максимального усилия. Утомляемость проверяется по методу эргографии. Для этого задается темп сжатий и их сила и измеряется работа, выполненная в пределах определённого интервала времени. В космическом полёте вместе с динамограммой регистрировалась ЭМГ длинного сгибателя предплечья.. Серебряные электроды с проводящей пастой закреплялись эластичными манжетами в верхней и нижней трети предплечья. Эта методика позволяет быстро определить наступление утомления: на динамограмме это проявляется в уменьшении амплитуды сжатий, а на ЭМГ – увеличением сигнала. Двигательная деятельность человека является основной формой его поведения во внешней среде. При этом следует указать, что не только физическая работа, но и разнообразные виды умственного труда в конечном итоге проявляются двигательной активностью. И. М. Сеченов (1863) по этому поводу писал, что «... все бесконечное разнообразие внешних проявлений мозговой деятельности сводится окончательно к одному лишь явлению — мышечному движению». . Основные принципы организации движений Выполнение двигательных актов осуществляется обширным комплексом нейронов, расположенных в различных отделах ЦНС. Такая функциональная система управления движениями является многоэтажной и многоуровневой. Общая схема управления движениями Решающим фактором поведения является полезный результат. Для его достижения в нервной системе формируется группа взаимосвязанных нейронов — функциональная система (П. К.Анохин, 1975). Деятельность ее включает следующие процессы: 1) обработка всех сигналов, поступающих из внешней и внутренней среды организма — так называемый афферентный синтез, 2) принятие решения о цели и задачах действия; 3) создание представления об ожидаемом результате и формирование конкретной программы движений; 4) анализ полученного результата и внесение в программу поправок — сенсорных коррекций. В процессах афферентного синтеза участвуют глубокие внутренние процессы — побуждение к действию (мотивация) и его замысел, извлекаются из памяти моторные следы (навыки) и выученные тактические комбинации. У человека на их основе создается определенный плани конкретная программа движения. Эти процессы отражаются в изменениях электрической активности мозга — «волна ожидания», изменения огибающей амплитуды ЭЭГ, усиление взаимосвязанности корковых нейронов, местные потенциалы готовности и др. феномены, связанные с повышением возбудимости корковых нейронов и созданием рабочей системы мозга. Выраженность этих феноменов отражает степень заинтересованности человека в реакции, скорость и силу ответных сокращений мышц. На уровне спинного мозга процессы преднастройки отражаются повышением возбудимости спиральных мотонейронов, в мышцах — повышением чувствительности проприорецепторов скелетных мышц. Сенсорная информация о результате выполнения движения, получаемая по каналам обратной связи, используется нервными центрами для уточнения временных, пространственных и силовых характеристик двигательных актов, внесения поправок в команды — так называемых сенсорных коррекций (W. к. Бернштейн, 1966; К. Прибрам, 1975). Выполнение двигательных актов осуществляется обширным комплексом нейронов, расположенных в различных отделах центральной нервной системы. Каждый из нервных центров, входящих в данный комплекс, может воздействовать как на нижележащие, так и на вышележащие центры. При этом между ними могут возникать циклы взаимных влияний от высших центров к низшим и обратно. Например, очень часто наблюдается циркуляция импульсов между корой и промежуточным мозгом: импульсы из коры больших полушарий могут влиять на активность нейронов промежуточного мозга, а те, в свою очередь, могут воздействовать на корковые нейроны. В каждом подобном цикле роль задающих звеньев принадлежит преимущественно вышележащим центрам, а активность нижележащих центров является регулируемым звеном. Эта активность поддерживается на заданном уровне в зависимости от потребностей организма. Нервная система действует по принципу рефлекса. Рефлекс - это реакция организма на раздражение, поступающее из внутренней или внешней среды, осуществляемая при посредстве ЦНС. Биологическая сущность рефлекса - в приспособлении организма к изменениям во внешней и внутренней среде. С помощью механизма рефлекса осуществляется единство организма и среды. Всякое мышечное движение имеет рефлекторную природу, рефлекторным же путем регулируется деятельность всех внутренних органов и систем. Рефлекторная дуга - путь, по которому распространяется возбуждение, - состоит из трех частей: афферентной (воспринимающей прибор и центростремительный нерв), центральной (участок ЦНС) и эфферентной (проводящей нервный путь к действующему органу). Эти части дуги могут быть простыми, состоящими из одной лишь нервной клетки, но могут быть и очень сложными, особенно центральная часть, которая может включать в себя много миллионов и даже сотен миллионов клеток (нейронов). Спортивная и трудовая деятельность человека, в том числе и овладение двигательными навыками, осуществляется по принципу взаимосвязи условных рефлексов и динамических стереотипов с безусловными рефлексами. Унаследованные рефлексы, от рождения заложенные в нервной системе, в ее структуре, в связях между нервными клетками, называют безусловными рефлексами. Объединяясь в длинные цепи, безусловные рефлексы являются основой инстинктивного поведения. У человека и у высших животных в основу поведения заложены условные рефлексы, вырабатываемые в процессе жизнедеятельности на основе безусловных рефлексов. Наличие циклических взаимосвязей между различными отделами нервной системы позволяет говорить о замкнутой системе регулирования, или замкнутом цикле регулирования. В процессе двигательной деятельности одновременно возникает множество замкнутых циклов регулирования (например, между корой и ретикулярной формацией, между корой и мозжечком, между мотонейронами спинного мозга и мышцей и др.). Таким образом, система регуляции движения является многоэтажной (многоуровневой) системой. Важную роль в этой сложной функциональной системе играет кора больших полушарий, которая управляет процессами, протекающими в этих циклах регулирования, и направляет их на решение общей задачи — достижение рабочего эффекта, или цели движения. Роль различных отделов цнс в регуляции позно-тонических реакций Мышечная деятельность включает в себя процессы осуществления двигательных актов и процессы поддержания позы тела. Эти процессы регулируются различными отделами ЦНС. Роль спинного мозга Мышечный тонус является по своей природе рефлекторным актом. Для его возникновения достаточна рефлекторная деятельность спинного мозга. При длительном растяжении мышц в поле силы тяжести возникает постоянное раздражение их проприорецепторов, потоки импульсов от которых проходят по толстым афферентным волокнам в спинной мозг, где передаются непосредственно (без участия вставочных нейронов) на альфа-мотонейроны передних рогов и вызывают тоническое напряжение мышц. Такие двухнейронные (или моносинаптические) рефлекторные дуги лежат в основе тонических сухожильных (с рецепторов сухожилий) и миотатических рефлексов на растяжение (с рецепторов мышечных веретен). Это рефлексы активного противодействия мышцы ее растяжению. В произвольной двигательной деятельности человека иногда требуется подавление этих рефлексов, например, при выполнении шпагата. Степень тонического напряжения мышцы зависит от частоты импульсов; посылаемых к ней альфа-мотонейронами. Однако, потоки этих импульсов могут регулироваться вышележащими этажами нервной системы, в частности, неспецифическими отделами ствола мозга с помощью так называемой гамма — регуляции. Разряды гамма-мотонейронов спинного мозга под влиянием ретикулярной формации повышают чувствительность рецепторов мышечных веретен. В результате при той же длине мышцы увеличивается поток импульсов от рецепторов. Роль коры головного мозга, мозжечка и ствола мозга В регуляции тонуса участвует также медленная часть пирамидной системы и различные структуры экстрапирамидной системы (подкорковые ядра, красные ядра и черная субстанция среднего мозга, мозжечок, ретикулярная формация ствола мозга, вестибулярные ядра продолговатого мозга). Неспецифическая система вызывает общее изменение тонуса различных мышц: усиление тонуса осуществляет активирующий отдел ретикулярной формации среднего мозга, а угнетение — тормозящий отдел продолговатого мозга. В отличие от нее специфическая система (моторные центры коры больших полушарий и ствола мозга) действует избирательно, т. е. на отдельные группы мышц-сгибателей или разгибателей. Усиление тонуса мышц-сгибателей вызывают корковые влияния, передающиеся непосредственно к нейронам спинного мозга по корково-спинномозговой (пирамидной) системе, а также через красные ядра (по корково-красноядерно-спинномозговой системе) и частично через ретикулярную формацию (по ретикуло-спинномозговой системе). В противоположность им, влияния, передающиеся через вестибулярные ядра продолговатого мозга к вставочным и двигательным нейронам передних рогов спинного мозга (по вестибуло-спинномозговым путям), вызывают тоническое повышение возбудимости мотонейронов мышц-разгибателей, что обеспечивает повышение тонуса этих мышц. Мозжечок формирует правильное распределение тонуса скелетных мышц: через красные ядра среднего мозга он повышает тонус мышц-сгибателей, а через вестибулярные ядра продолговатого мозга — усиливает тонус мышц-разгибателей. В поддержании позы и равновесия тела, регуляции тонуса мышц основное значение имеет медиальная продольная зона мозжечка — кора червя. При мозжечковых расстройствах падает тоническое напряжение мышц (атония) и вследствие ненормального распределения тонуса мышц конечностей возникает нарушение походки (атаксия). Бледное ядро угнетает тонус мышц, а полосатое тело снижает его угнетающее действие. Высший контроль тонической активности мышц осуществляет кора больших полушарий, в частности ее моторные, премоторные и лобные области. С ее участием происходит выбор наиболее целесообразной для данного момента позы тела, обеспечивается ее соответствие двигательной задаче. Непосредственное отношение к регуляции тонуса мышц имеют медленные пирамидные нейроны положения. Корковые влияния натонические реакции мышц передаются через медленную часть пирамидного тракта и через экстрапирамидную систему. Альфа-мотонейронам и далее к мышце, повышая ее тонус. |