1. 1Роль физиологии в материалистическом понимании сущности жизни. Значение работ И. М. Сеченова и И. П. Павлова в создании материалистических основ физиологии
 Скачать 3.27 Mb. Скачать 3.27 Mb.
|

|
28.17 Рецепторы органов чувств, понятие, классификация , основные свойства и особенности. Механизм возбуждения. Понятие функциональной мобильности. Рецептором называют генетически детерменированные макромолекулы, которые располагаются на субсинаптической мембране и взаимодействуют с молекулами медиатора как ключ с замком, запуская при этом рективную систему синапса. Рецепторы делят на четыре группы: механо-, термо-, хемо- и фоторецепторы. Каждую группу подразделяют на более узкие диапазоны рецепции. Например, зрительные рецепторы делятся на воспринимающие освещенность, цвет, слуховые — определенный тон, вкусовые — определенные вкусовые раздражения (соленое, сладкое, горькое) и т. д. Рецепторный потенциал возникает при раздражении рецептора как результат деполяризации и повышения проводимости участка его мембраны, который называется рецептивным. Рецептивный участок мембраны имеет специфические свойства, в том числе биохимические, отличающие его от мембраны тела и аксона. Возникший в рецептивных участках мембраны рецепторный потенциал электротонически распространяется на аксонный холмик рецепторного нейрона, где возникает генераторный потенциал. Возникновение генераторного потенциала в области аксонного холмика объясняется тем, что этот участок нейрона имеет более низкие пороги возбуждения и потенциал действия в нем развивается раньше, чем в других частях мембраны нейрона. Чем выше генераторный потенциал, тем интенсивнее частота разрядов распространяющегося потенциала действия от аксона к другим отделам нервной системы. Следовательно, частота разрядов рецепторного нейрона зависит от амплитуды генераторного потенциала. Рецепторные нейроны различаются по скорости уменьшения их реакции (адаптации) на длящуюся стимуляцию. Рецепторные нейроны, медленно адаптирующиеся к раздражению, т. е. длительное время генерирующие потенциалы действия, называются тоническими. Рецепторы, быстро и коротко реагирующие на стимуляцию группой импульсов, называются физическими. Таким образом, реакция рецепторного нейрона, предназначенного для передачи информации из области восприятия, имеет 5 стадий: 1) преобразование сигнала внешнего раздражения; 2) генерация рецепторного потенциала; 3) распространение рецепторного потенциала по нейрону; 4) возникновение генераторного потенциала; 5) генерация нервного импульса. Мобильность функциональная — непрерывное изменение числа и возбудимости работающих рецепторов, обусловленное в основном влиянием на них со стороны ц.н.с. 29.1 нейрон как структурно-функциональная единица в ЦНС. Классификация нейронов по структурным и функциональным признакам. Механизм проникновения возбуджения в нейроне. Интегративная функция нейрона. Нейроны — специализированные клетки, способные принимать, обрабатывать, кодировать, передавать и хранить информацию, организовывать реакции на раздражения, устанавливать контакты с другими нейронами, клетками органов. Уникальными особенностями нейрона являются способность генерировать электрические разряды и передавать информацию с помощью специализированных окончаний — синапсов. Функционально в нейроне выделяют следующие части: воспринимающую — дендриты, мембрана сомы нейрона; интегративную — сома с аксонным холмиком; передающую — аксонный холмик с аксоном. Тело нейрона (сома) выполняет информационную, трофическую функцию, также рост дендритов и аксона. Мембрана из 2-ух слоев липидных молекул. Имеются Рибосомы, Базофильное вещество, аппарат Гольджи, Лизосомы, Митохондрии, Ядро, ядрышко. Дендриты — основное воспринимающее поле нейрона. Аксон представляет собой вырост цитоплазмы, приспособленный для проведения информации, собранной дендритами, переработанной в нейроне и переданной аксону. По строению нейроны делят на три типа: униполярные, биполярные и мультиполярные. Истинно униполярные нейроны находятся только в мезэнцефалическом ядре тройничного нерва. Эти нейроны обеспечивают проприоцептивную чувствительность жевательных мышц. Другие униполярные нейроны называют псевдоуниполярными, на самом деле они имеют два отростка,и все эти клетки располагаются в сенсорных узлах: спинальных, тройничном и т. д. Они обеспечивают восприятие болевой, температурной, тактильной, проприоцептивной, бароцептивной, вибрационной сигнализации. Биполярные нейроны имеют один аксон и один дендрит. Нейроны этого типа встречаются в основном в периферических частях зрительной, слуховой и обонятельной систем. Биполярные нейроны дендритом связаны с рецептором, аксоном — с нейроном следующего уровня организации соответствующей сенсорной системы. Мультиполярные нейроны имеют несколько дендритов и один аксон. Существует классификация нейронов, учитывающая химическую структуру выделяемых в окончаниях их аксонов веществ: холинергические, пептидергические, норадреналинергические, дофаминергические, серотонинергические и др. По чувствительности к действию раздражителей нейроны делят на моно-, би-, полисенсорные. По положению в рефлекторной дуге различают афферентные нейроны (чувствительные нейроны), эфферентные нейроны (двигательными нейронами) и интернейроны (вставочные нейроны) Афферентные нейроны – проводят импульс от поверхности тела и внутренних органов в мозг. Эфферентные нейроны– проводят импульс от головного и спинного мозга ко всем рабочим органам. Вставочные нейроны – анализируют информацию и вырабатывают решения. Интегративная функция нейрона. Общее изменение мембранного потенциала нейрона является результатом сложного взаимодействия (интеграции) местных ВозбПСП и ТормПСП всех многочисленных активированных синапсов на теле и дендритах клетки. На мембране нейрона происходит процесс алгебраического суммирования положительных и отрицательных колебаний потенциала. При одновременной активации нескольких возбуждающих синапсов общий ВПСП нейрона представляет собой сумму отдельных местных ВПСП каждого синапса. При одновременном возникновении двух различных синаптических влияний — ВПСП и ТПСП — происходит взаимное вычитание их эффектов. В конечном итоге реакция нервной клетки определяется суммой всех синаптических влияний. Преобладание тормозных синаптических воздействий приводит к гиперполяризации мембраны и торможению деятельности клетки. При сдвиге мембранного потенциала в сторону деполяризации повышается возбудимость клетки. Ответный разряд нейрона возникает лишь тогда, когда изменения мембранного потенциала достигают порогового значения — критического уровня деполяризации. Механизм возникновения возбуждения.На дендритах нейронов большей частью формируются возбуждающие синапсы, а на теле клетки – тормозные. Если возбуждающие и тормозные синапсы активируются одновременно, то возникающие локальные токи электротонически достигают аксонного холмика и там суммируются. Суммация не является линейной. В случае изменения мембранного потенциала аксонного холмика до критического уровня деполяризации на аксоне возникает потенциал действия (процесс возбуждения). Если же в процессе суммации возбуждающих и тормозных постсинаптических токов сдвиг мембранного потенциала оказался ниже критического, то потенциал действия не образуется; нейрон будет заторможен. Вопрос 30.2 Определение нервного центра (классическое и современное). Свойства нервных центров, обусловленные их структурными звеньями (иррадация, конвергенция, последействием возбуждения) Нервный центр- это динамическая совокупность взаимосвязанных нейронов, расположенных в разных отделах ЦНС, которая формируется в процессе развития организма и обеспечивать какую-либо функцию. Нервный центр-это совокупность взаимосвязанных нейронов, расположенных в какой-либо области ЦНС, которая обеспечивает рефлекторных ответ. Свойства НЦ: 1.чувствительность к гипоксии 2.чувствительность к конц. Глюкозы в крови 3. Односторонность проведения возбуждения 4. Иррадиация возбуждения. Иррадация - распределение возбуждения от одного к множеству. В естественных условиях ее не бывает. Возбуждение неограниченно распространяется. 5. Суммация возбуждения. 6. Наличие синаптической задержки. Время рефлекторной реакции зависит в основном от двух факторов: скорости движения возбуждения по нервным проводникам и времени распространения возбуждения с одной клетки на другую через синапс. При относительно высокой скорости распространения импульса по нервному проводнику основное время рефлекса приходится на синаптическую передачу возбуждения (синаптическая задержка). 7.Высокая утомляемость. Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомлением 8. Тонус. Тонус, определяется тем, что в покое в отсутствие специальных внешних раздражений определенное количество нервных клеток находится в состоянии постоянного возбуждения, генерирует фоновые импульсные потоки 9. Пластичность. 10. Конвергенция. Количественное соотношение периферических рецепторных и промежуточных центральных нейронов (10:1) предполагает значительную конвергенцию («сходимость») разномодальных сенсорных посылок на одни и те же центральные нейроны. 11. Интеграция в нервных центрах. Важные интегративные функции клеток нервных центров ассоциируются с интегративными процессами на системном уровне в плане образования функциональных объединений отдельных нервных центров в целях осуществления сложных координированных приспособительных целостных реакций организма (сложные адаптивные поведенческие акты). 12. Свойство доминанты. Доминантным называется временно господствующий в нервных центрах очаг (или доминантный центр) повышенной возбудимости в центральной нервной системе. По А.А.Ухтомскому, доминантный нервный очаг характеризуется такими свойствами, как повышенная возбудимость, стойкость и инертность возбуждения, способность к суммированию возбуждения. 13. Цефализация нервной системы. Основная тенденция в эволюционном развитии нервной системы проявляется в перемещении, сосредоточении функции регуляции и координации деятельности организма в головных отделах ЦНС. Этот процесс называется цефализацией управляющей функции нервной системы. Вопрос 31.3 Физиологические свойства нервных центров: пространственная и временная суммация, трансформация ритма, посттетаническая потенциация, низкая лабильность, утомляемость, чувствительность к нейротропным веществам, автоматия. 1. Пространственная и временная суммация. Временная суммация возникает, как и в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда ВПСП. Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторов нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются и в мембране нейрона генерируется распространяющийся ПД. 2. Трансформация ритма возбуждения – изменение частоты нервных импульсов при прохождении через нервный центр. Частота может понижаться или повышаться. Например, повышающая трансформация (увеличение частоты) обусловлено дисперсией и мультипликацией возбуждения в нейронах. Первое явление возникает в результате разделения нервных импульсов на несколько нейронов, аксоны которых образуют затем синапсы на одном нейроне (рис). Второе, генерацией нескольких нервных импульсов при развитии возбуждающего постсинаптического потенциала на мембране одного нейрона. Понижающая трансформация объясняется суммацией нескольких ВПСП и возникновением одного ПД в нейроне. 3. Посттетаническая потенциация, это усиление рефлекторной реакции в результате длительного возбуждения нейронов центра. Под влиянием многих серий нервных импульсов, проходящих с большой частотой через синапсы,. выделяется большое количество нейромедиатора в межнейронных синапсах. Это приводит к прогрессирующему нарастанию амплитуды возбуждающего постсинаптического потенциала и длительному (несколько часов) возбуждению нейронов. 4. Низкая физиологическая лабильность и быстрая утомляемость. Н.Ц. могут проводить импульсы лишь ограниченной частоты. Их утомление объясняется утомлением синапсов и ухудшением метаболизма нейронов. 6. Автоматия или спонтанная активность нервных центров. Периодическая или постоянная генерация нейронами нервных импульсов, которые возникают в них самопроизвольно, т.е. в отсутствии сигналов от других нейронов или рецепторов. Обусловлена колебаниями процессов метаболизма в нейронах и действием на них гуморальных факторов. Вопрос 32.4 Торможение в ЦНС ( И.М. Сеченов). Современные представления об основных видах центрального торможения постсинаптического, пресинаптического и их механизмах. Торможение – активный, срочный процесс прекращения или задержки текущей функции под действием раздражителя. Процесс торможения в центральной нервной системе был открыт в 1862 г. И. М. Сеченовым. В опытах на лягушках он делал поперечные разрезы головного мозга на различных уровнях и раздражал нервные центры, накладывая на разрез кристаллик поваренной соли. При этом обнаруживалось, что при раздражении промежуточного мозга наступает угнетение или полное торможение спинномозговых рефлексов: лапка лягушки, погруженная в слабый раствор серной кислоты, не отдергивалась. Постсиналтическое торможение — основной вид торможения, развивающийся в постсинаптической мембране аксосоматических и аксодендритических синапсов под влиянием активации тормозных нейронов, в концевых разветвлениях аксонных отростков которых освобождается и поступает в синаптическую щель тормозной медиатор. Тормозной эффект таких нейронов обусловливается специфической природой медиатора — химического переносчика сигнала с одной клетки на другую. Наиболее распространенным тормозным медиатором является гамма-аминомасляная кислота (ГАМК). Химическое действие ГАМК вызывает в постсинаптической мембране эффект гиперполяризации в виде тормозных постсинаптических потенциалов (ТПСП), пространственно-временная суммация которых повышает уровень мембранного потенциала (гиперполяризация), приводит к урежению или полному прекращению генерации распространяющихся ПД. 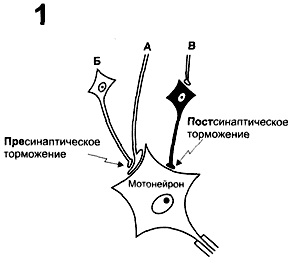 А – возбуждающее окончание;Б – нейрон, вызывающий пресинаптическое торможение; В – нейрон, вызывающий постсинаптическое торможение. А – возбуждающее окончание;Б – нейрон, вызывающий пресинаптическое торможение; В – нейрон, вызывающий постсинаптическое торможение.Пресинаптическое торможение возникает перед синаптическим контактом – в пресинаптической области. Окончание аксона одной нервной клетки (терминаль) образует синапс на окончании аксона другой нервной клетки и блокирует передачу возбуждения в последней. В области такого пресинаптического контакта развивается чрезмерно сильная деполяризация мембраны аксона, которая приводит к угнетению проходящих здесь потенциалов действия. Пресинаптическое торможение осуществляют нейроны (Б), аксоны которых оканчиваются на возбуждающих синаптических окончаниях другого нейрона (А), образуя аксо-аксональные синапсы (рис. 1Б). Пресинаптическое торможение выполняется посредством одного из двух механизмов. 1. Пресинаптический тормозным нейрон Б выделяет нейромедиатор, который увеличивает Cl–проводимость и вызывает гиперполяризацию мембраны возбуждающего нервного окончания А. Это приводит к снижению возбудимости и увеличению порога генерации ПД возбуждающего окончания. 2. Пресинаптический возбуждающий нейрон выделяет нейромедиатор, вызывающий длительную деполяризацию мембраны, что приводит к инактивации потенциалозависимых Na+-каналов нервного окончания и, соответственно, торможению его активности. Вопрос 33.5 Определение координации в ЦНС. Основные принципы координационной деятельности ЦНС: рецепрокность, общего «конечного» пути, доминанты, временной связи, обратной связи. Координация - процессы взаимодействия между нейронами и нервным центром, которое обеспечивает их согласованную деятельность. Принцип доминанты. «… господствующий очаг возбуждения в ЦНС, который определяет текущую деятельность или рабочую позу организма, я обозначил термином доминанта» -Ухтомский. Свойство доминанта:1. повышенная возбудимость2. стойкость доминанты: могут сохранять часы, месяца, годы, всю жизнь 3.Инерция доминанты –последствие. 4. Способность к суммации Сущность его заключается в том, что среди множества возбужденных очагов в ЦНС, существует очаг господствующего возбуждения. Этот доминантный очаг подчиняет себе все другие очаги возбуждения. Принцип обратной связи. Всякий двигательный акт, вызванный афферентным раздражителем, сопровождается возбуждением рецепторов мышц, сухожилий, суставных сумок. Эти сигналы с проприорецепторов поступают в ЦНС. Обратная связь позволяет соотнести выраженность изменений параметров системы с ее работой. Связь выхода системы с ее входом с положительным коэффициентом усиления называется положительной обратной связью, а с отрицательным коэффициентом — отрицательной обратной связью. Положительная обратная связь в основном характерна для патологических ситуаций. Отрицательная обратная связь обеспечивает устойчивость системы. Механизмы обратной связи обеспечивают поддержание всех констант гомеостаза. Принцип реципрокности. Он отражает характер отношение между центрами, ответственными за осуществление противоположных функций (вдоха и выдоха, сгибания и разгибания конечностей), и заключается в том, что нейроны одного центра, возбуждаясь, тормозят нейроны другого и наоборот. Принцип общего конечного пути. Импульсы, приходящие в ЦНС по разным афферентным волокнам, могут сходиться к одним и тем же вставочным, или эфферентным, нейронам. Шеррингтон назвал это явление «принципом общего конечного пути». Один и тот же мотонейрон может возбуждаться импульсами, приходящими от различных рецепторов (зрительных, слуховых, тактильных), т.е. участвовать во многих рефлекторных реакциях. Принцип временной связи. Высшая форма временной связи - условный рефлекс. |