1. 1Роль физиологии в материалистическом понимании сущности жизни. Значение работ И. М. Сеченова и И. П. Павлова в создании материалистических основ физиологии
 Скачать 3.27 Mb. Скачать 3.27 Mb.
|

|
13.2 Законы раздражения возбудимых тканей: значение силы раздражителя, частоты раздражителя, его длительности, крутизны его нарастания. 1. Постоянный ток раздражает только при своем изменении 2. Раздражающие действия постоянного тока тем сильнее, чем быстрее он меняется во времени(чем выше скорость нарастания), значит макс. эффективность у мгновенного тока (Р-импульсы) 3. Катод повышает возбудимость и возбуждает, анод снижает возбудимость и уменьшает текущее возбуждение(тормозит). Закон полярного действия постоянного тока ( 1 закон Пфлюгера) – закон о месте и времени возникновения возбуждения в клетке: При действии на ткань постоянного тока возбуждение под катодом возникает в момент замыкания цепи, а под анодом в момент размыкания. Раздражающее действие катода выражено сильнее, чем анода. 2. Закон физиологического электротона (2 закон Пфлюгера) – закон об изменении физиологических свойств тканей при прохождении через них постоянного тока: При прохождении через ткань постоянного тока в области катода возбудимость и проводимость повышаются, а лабильность снижается(физиологический Кат-электротон), в области анода возбудимость и проводимость понижается, а лабильность повышается – физиологический АН-электротон. Дополнение Вериго – при прохождении через ткань сильного или длительного постоянного тока повышенная возбудимость в области катода сменяется пониженной – катодическая депрессия, а пониженная возбудимость в области анода сменяется повышенной – анодическая экзальтация. ИТОГ: Возбудимость клетки не зависит ни от величины мембранного потенциала, ни от величины критического уровня деполяризации, а определяется их соотношением: чем меньше между ними разница, тем выше возбудимость и наоборот Закон Дюбуа-Раймона: Раздражающее действие постоянного тока не зависит ни от силы тока (его плотности), ни от длительности действия(времени), а определяется скоростью его нарастания: чем быстрее меняется ток во времени, тем большей раздражающей силой он обладает. Закон «Кривая Сила-Время»: Раздражающее действие постоянного тока тем сильнее, чем больше время его действия Хроноксия – время, в теч которого на ткань должен действовать постоянный ток, равным 2 реобазам, чтобы вызвать возбуждение. 14.3 Современные представления о строении и функции мембран. Ионные каналы мембран. Ионные градиенты клетки, механизмы из возникновения. Функции: 1. Барьерная – мембрана при помощи соответствующих механизмов участвует в создании концентрационных градиентов, препятствуя свободной диффузии. 2.Регуляторная функция клеточной мембраны заключается в тонкой регуляции внутриклеточного содержимого и внутриклеточных реакций за счет рецепции внеклеточных биологически активных веществ, что приводит к изменению активности ферментных систем мембраны и запуску механизмов вторичных «месенджеров» («посредников»). 3.Преобразование внешних стимулов неэлектрической природы в электрические сигналы (в рецепторах). 4.Высвобождение нейромедиаторов в синаптических окончаниях. Жидкостно-мозаичная модель Сингера и Николсона: В фосфолипидном бислое интегрированы глобулярные белки, полярные участки которых образуют гидрофильную поверхность в водной фазе. Эти интегрированные белки выполняют различные функции, в том числе рецепторную, ферментативную, образуют ионные каналы, являются мембранными насосами и переносчиками ионов и молекул. Некоторые белковые молекулы свободно диффундируют в плоскости липидного слоя; в обычном состоянии части белковых молекул, выходящие по разные стороны клеточной мембраны, не изменяют своего положения. Особая морфология клеточных мембран определяет их электрические характеристики, среди которых наиболее важными являются емкость и проводимость. Емкостные свойства в основном определяются фосфолипидным бислоем, который непроницаем для гидратированных ионов и в то же время достаточно тонок (около 5 нм), чтобы обеспечивать эффективное разделение и накопление зарядов и электростатическое взаимодействие катионов и анионов Проводимость (g) — величина, обратная электрическому сопротивлению и равная отношению величины общего трансмембранного тока для данного иона к величине, обусловившей его трансмембранной разности потенциалов. Строение и функции ионных каналов. Ионы Na+, K+, Са2+, Сl- проникают внутрь клетки и выходят наружу через специальные, заполненные жидкостью каналы. Размер каналов довольно мал (диаметр 0,5—0,7 нм). Рассмотрим принцип работы ионных каналов на примере натриевого канала. Полагают, что в состоянии покоя натриевый канал закрыт. При деполяризации клеточной мембраны до определенного уровня происходит открытие m-активационных ворот (активация) и усиление поступления ионов Na+ внутрь клетки. Через несколько миллисекунд после открытия m-ворот происходит закрытие п-ворот, расположенных у выхода натриевых каналов (инактивация) (рис. 2.4). Инактивация развивается в клеточной мембране очень быстро и степень инактивации зависит от величины и времени действия деполяризующего стимула. При генерации одиночного потенциала действия в толстом нервном волокне изменение концентрации ионов Na+ во внутренней среде составляет всего 1/100000 от внутреннего содержания ионов Na гигантского аксона кальмара. Однако для тонких нервных волокон это изменение концентрации может быть весьма существенным. Кроме натриевых, в клеточных мембранах установлены другие виды каналов, избирательно проницаемых для отдельных ионов: К+, Са2+ Ходжкин и Хаксли сформулировали принцип «независимости» каналов, согласно которому потоки натрия и калия через мембрану независимы друг от друга. Свойство проводимости различных каналов неодинаково. В частности, для калиевых каналов процесс инактивации, как для натриевых каналов, не существует. Имеются особые калиевые каналы, активирующиеся при повышении внутриклеточной концентрации кальция и деполяризации клеточной мембраны. Активация калий-кальцийзависимых каналов ускоряет реполяризацию, тем самым восстанавливая исходное значение потенциала покоя. Особый интерес представляют кальциевые каналы. Входящий кальциевый ток, как правило, недостаточно велик, чтобы нормально деполяризовать клеточную мембрану. Чаще всего поступающий в клетку кальций выступает в роли вторичного посредника. Активация кальциевых каналов обеспечивается деполяризацией клеточной мембраны, например входящим натриевым током. Процесс инактивации кальциевых каналов достаточно сложен. С одной стороны, повышение внутриклеточной концентрации свободного кальция приводит к инактивации кальциевых каналов. С другой стороны, белки цитоплазмы клеток связывают кальций, что позволяет поддерживать длительное время стабильную величину кальциевого тока, хотя и на низком уровне; при этом натриевый ток полностью подавляется. Кальциевые каналы играют существенную роль в клетках сердца. 15.4 Мембранный потенциал, теория его происхождения. Мембранный потенциал - это разность потенциалов между наружной и внутренней поверхностями элементарной пограничной мембраны клетки Мембранный потенциал – сила электростатического взаимодействия, которая уравновешивает электрические токи, протекающие через мембрану изнутри кнаружи, снаружи – внутрь. Современная теория Ходжкина-Хаксли-Катца: 1. В основе возникновения биопотенциала является обмен веществ и энергии между клеткой и окружающей средой 2. Местом возникновения биопотенциала является поверхностная мембрана клетки 3. Непосредственной причиной возникновения биопотенциала является разность концентрации ионов (ионная асимметрия вне и внутри клетки) 4. Поверхностная пограничная мембрана клетки обладает свойством полупроницаемости, которое может меняться при различных состояниях. 16.5. Потенциал действия, его фазы. Динамика проницаемости мембраны в различные фазы потенциала действия. Под потенциалом действия понимают быстрое колебание потенциала, сопровождающееся, как правило, перезарядкой мембраны. Потенциал действия – это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны. При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя – положительный. Компоненты потенциала действия: 1) локальный ответ; 2) высоковольтный пиковый потенциал (спайк); 3) следовые колебания. Ионы Na путем простой диффузии поступают в клетку без затрат энергии. Достигнув пороговой силы, мембранный потенциал снижается до критического уровня деполяризации (примерно 50 мВ). Критический уровень деполяризации – это то количество милливольт, на которое должен снизиться мембранный потенциал, чтобы возник лавинообразный ход ионов Na в клетку. Высоковольтный пиковый потенциал (спайк). Пик потенциала действия является постоянным компонентом потенциала действия. Он состоит из двух фаз: 1) восходящей части – фазы деполяризации; 2) нисходящей части – фазы реполяризации. Быстрое и большое количество поступление ионов Na в клетку приводит к изменению потенциала на клеточной мембране. Чем больше ионов Na войдет в клетку, тем в большей степени деполяризуется мембрана, тем больше откроется активационных ворот. Возникновение заряда с противоположным знаком называется инверсией потенциала мембраны. Движение ионов Na внутрь клетки продолжается до момента электрохимического равновесия по иону Na Амплитуда потенциала действия не зависит от силы раздражителя, она зависит от концентрации ионов Na и от степени проницаемости мембраны к ионам Na. Нисходящая фаза (фаза реполяризации) возвращает заряд мембраны к исходному знаку. При достижении электрохимического равновесия по ионам Na происходит инактивация активационных ворот, снижается проницаемость к ионам Na и возрастает проницаемость к ионам K. Полного восстановления мембранного потенциала не происходит. В процессе восстановительных реакций на клеточной мембране регистрируются следовые потенциалы – положительный и отрицательный. 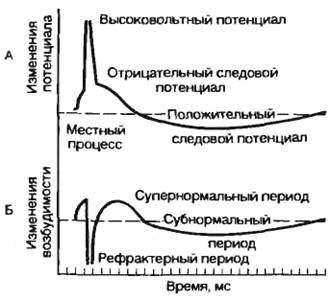 17.6 Возбудимость, методы её оценки. Изменения возбудимости при действии постоянного тока (электротон, катодическая депрессия, аккомодация). Возбудимость – это способность возбудимых тканей на действие раздражителя отвечать возбуждением, которое проявляется в виде биоэлектрического процесса и специфической ответной реакции. Подпороговое пассивное поведение клеточной мембраны называется электротоническим, или электротоном. Закон физиологического электротона (2 закон Пфлюгера) – закон об изменении физиологических свойств тканей при прохождении через них постоянного тока: При прохождении через ткань постоянного тока в области катода возбудимость и проводимость повышаются, а лабильность снижается(физиологический Кат-электротон), в области анода возбудимость и проводимость понижается, а лабильность повышается – физиологический АН-электротон. Дополнение Вериго – при прохождении через ткань сильного или длительного постоянного тока повышенная возбудимость в области катода сменяется пониженной – катодическая депрессия, а пониженная возбудимость в области анода сменяется повышенной – анодическая экзальтация. ИТОГ: Возбудимость клетки не зависит ни от величины мембранного потенциала, ни от величины критического уровня деполяризации, а определяется их соотношением: чем меньше между ними разница, тем выше возбудимость и наоборот Закон Дюбуа-Раймона: Раздражающее действие постоянного тока не зависит ни от силы тока (его плотности), ни от длительности действия(времени), а определяется скоростью его нарастания: чем быстрее меняется ток во времени, тем большей раздражающей силой он обладает. Аккомодация ткани – явление приспособления (снижения возбудимости) ткани к действию медленно-нарастающего раздражителя. 18.7 Соотношения фаз изменения возбудимости при возбуждении с фазами потенциала действия. 1) местный ответ - физиологический катэлектротон. 2) высоковольтный пик - катодическая депрессия 3) следовая деполяризация - катэлектротон 4) следовая гиперполяризация - анэлектротон Когда ткань возбуждается генерирует ПД, то временно в ней меняется возбудимость: сначала ткань становится совершенно невозбудимой (абсолютная рефрактерность) любой по силе стимул не способен вызывать в ней новый приступ возбуждения, эта фаза обычно наблюдается во время пика ПД, затем происходит постепенное восстановление возбудимости до исходного состояния (фаза относительной рефрактерности) – в этот момент раздражитель может вызвать возбуждение (генерацию нового ПД), но для этого он должен быть выше порогового (исходного). Затем в фазу следовой негативности возбудимость повышается (супервозбудимость или фаза экзальтации). В этот момент подпороговые раздражители могут вызвать возбуждение. В тканях, в которых ярко проявилась следовая гиперполяризация наблюдается еще одна фаза- субнормальности (фаза сниженной возбудимости). 19.8 Строение и классификация синапсов. Механизм передачи сигналов в синапсах ( электрических и химических) Ионные механизмы постсинаптических потенциалов, их виды.Синапсом называют образование, обеспечивающее передачу возбуждения от одной структуры к другой. Синапсы по способу передачи сигнала: I.Химические (передача сигналов с помощью медиатора) II.Электрические (нексусы) – нет у высш животных и у человека Синапсы: I. Центральные (>60) II. Периферические : 1) Соматические (нервно-мышечные) 2) Вегетативные: 1. Ганглионарные 2. Эффекторные(симпатические и парасимпатические) Синапсы по характеру действия на воспринимающую структуру: I. возбуждающие II.тормозящие По местоположению выделяют нервно-мышечные синапсы и нейронейрональные, последние в свою очередь делятся на аксосоматические, аксоаксональные, аксодендритические, дендросоматические. В химическом синапсе выделяют 3 элемента : пресинаптическую область (пресинаптическая терминаль); синаптическую щель; постсинаптическую область. Пресинаптическая область представляет собой демиелинизированный терминальный участок отростка нервной клетки. По форме пресинаптическая терминаль напоминает собой колбу, прилежащую основанием к участку мембраны возбудимой клетки. Наиболее существенной чертой пресинаптической области является скопление пресинаптических пузырьков диаметром 50 мм (везикул), в которых содержится медиатор (химическое соединение, которое является материальным носителем сигнала к реципиентской клетке). Кроме везикул с медиатора в пресинаптической области содержится большое количество митохондрий и лизосом, что свидетельствует о высокой активности обменных процессов в этой области. Кроме того, в этой области обнаружены предшественники медиаторов и продукты их метаболизма. Синаптическая щель. В химических синапсах составляет от 20 до 50 мм. Здесь содержаться вода, электролиты, олигосахариды, ферменты, участвующие в расщеплении медиатора. Постсинаптическая область. Включает субсинаптическую мембрану (участок постсинаптической мембраны, имеющий специальный аппарат -рецепторы, характеризующиеся сродством к медиатору. В этой же области имеются химически-чувствительные ионные каналы. Собственно постсинап-тическая мембрана - участок постсинаптической мембраны, которая содержит потенциал-зависимые ионные каналы и на которой происходит генерация постсинаптических потенциалов. Современные представления о механизмах передачи возбуждения в синапсе. ПД распространяется по нервному волокну к пресинаптической области. Изменение проницаемости мембраны пресинаптического образования к ионам Са++ и поступление Са++ в пресинаптическом образовании. Движение везикул с активным медиатором пресинаптической области к пресинаптической мембране и выделение медиатора в синаптическую щель методом экзоцитоза. Движение медиатора к субсинаптической мембране постсинаптической области и взаимодействие с соответствующими рецепторами мембраны. Далее возможны два варианта: 1- либо само взаимодействие медиатора с рецептором приводит к активации ионных каналов субсинаптической мембраны (это имеет место в некоторых холинэргических синапсах; 2 - либо вначале активируется вторичный посредник, а затем уже включается цепь биохимических процессов, приводящая к изменению проницаемости ионных каналов. Изменение проницаемости ионных каналов приводит к формированию местных токов и генерации на постсинаптической мембране постсинаптического потенциала. В зависимости от того по отношению к какому иону меняется проницаемость мембран, возможны два варианта постсинаптических потенциалов - возбуждающий, деполяризационный (ВПСП) и тормозной, гиперполяризационный (ТПСП). В основе ВПСП лежит повышение проницаемости мембраны по отношению к ионам Nа+, а ТПСП- по отношению к ионам К+ и Cl- . Характер активности тех или иных каналов определяется химической структурой медиатора, особенностью рецепторного образования, а также связанного с ним вторичного посредника. Электрические синапсы обладают односторонним проведением возбуждения. Это легко доказать при регистрировании электрического потенциала на синапсе: при раздражении афферентных путей мембрана синапса деполяризуется, а при раздражении эфферентных волокон — гиперполяризуется. Оказалось, что синапсы нейронов с одинаковой функцией обладают двусторонним проведением возбуждения (например, синапсы между двумя чувствительными клетками), а синапсы между разнофункциональными нейронами (сенсорные и моторные) обладают односторонним проведением. Функции электрических синапсов заключаются прежде всего в обеспечении срочных реакций организма. Этим, видимо, объясняется расположение их у животных в структурах, обеспечивающих реакцию бегства, спасения от опасности и т. д. Электрический синапс сравнительно мало утомляем, устойчив к изменениям внешней и внутренней среды. Видимо, эти качества наряду с быстродействием обеспечивают высокую надежность его работы. |