Тема 2. ЗАГАЛЬНА ФІЗІОЛОГІЯ РЕГУЛЯЦІЇ ФУНКЦІЙ ОРГАНІЗМУ. 1. Біологічна регуляція, її види і значення. Контур біологічної регуляції. Роль зворотнього звязку в регуляції
 Скачать 3.29 Mb. Скачать 3.29 Mb.
|

|
загальна фізіологія регуляції функцій організму. 1. Біологічна регуляція, її види і значення. Контур біологічної регуляції. Роль зворотнього зв’язку в регуляції. Біологічна регуляція – процес взаємодії елементів організму, спрямований на отримання пристосувального (корисного) результату. Біологічна регуляція   Внутрішньоклітинна Організмова     НЕРВОВА ГУМОРАЛЬНА ГЕННА ФЕРМЕНТАТИВНА МІОГЕННА Елементи організму, які взаємодіють при виконанні акта біологічної регуляції, можна зобразити у вигляді окремих блоків та об’єднати у вигляді контуру біологічної регуляції. Т 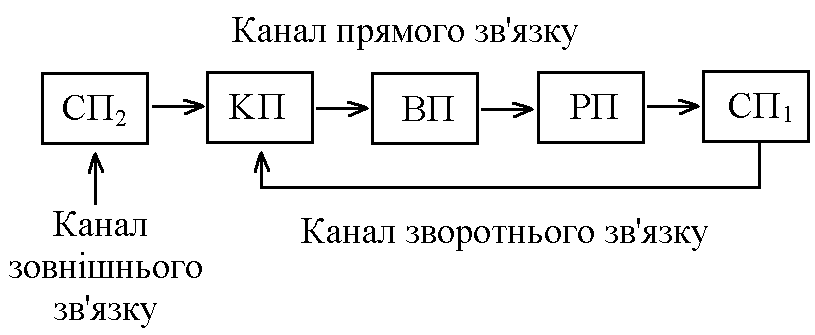 аким чином в контурі біологічної регуляції можна виділити наступні елементи: аким чином в контурі біологічної регуляції можна виділити наступні елементи:1. РП – регульований параметр, котрий завдяки біологічній регуляції: - підтримується на необхідному рівні; - змінюється в необхідному напрямку. 2. СП – слідкуючий пристрій (рецептори), що реєструють величину РП. 3. КП – керуючий пристрій – нервовий центр чи ендокринна залоза, що на основі аналізу інформації про величину РП, про її зміни, виробляє керуючий сигнал (при нервовій регуляції – серія ПД, при гуморальній – гормон) і посилає її до органів-ефекторів. Нервовий центр як керуючий пристрій має такі структурні елементи: - уставка – зберігає інформацію про необхідну величину РП; - блок порівняння – порівнює інформацію про задану величину РП з її реальною величиною, тобто встановлює наявність відхилень РП; - блок управління – з урахуванням наявності чи відсутності відхилення РП від заданого рівня виробляє керуючий сигнал і посилає його до органів-ефекторів. 4. ВП – виконавчий пристрій – органи-ефектори, які під впливом керуючого сигналу змінюють свою діяльність таким чином, що РП підтримується на заданому рівні чи змінюється в необхідному напрямку. Перелічені вище елементи контура біологічної регуляції зв’язуються між собою каналами зв’язку, якими в контурі передається інформація: - канал прямого зв’язку – по ньому передається керуючий сигнал від КП до ВП підтримка заданого рівня або зміна РП; - канал зворотнього зв’язку – по ньому передається інформація з СП1 в КП: - про поточну величину РП; - про ефективність керуючих сигналів, що вироблені КП і спрямовані на усунення відхилення РП від заданого рівня або на його зміну в потрібному напрямку. КП, ВП, РП, СП1 за допомогою каналів прямого та зворотного зв’язку утворюють замкнутий контур регуляції, котрий забезпечує здійснення регуляції “за відхиленням” (саморегуляції на основі негативного зворотнього зв’язку). Ця саморегуляція здійснюється наступним чином: певні чинники викликають зміни РП (його відхилення від заданого рівня) інформація про це сприймається рецепторами (СП1) та по каналу зворотнього зв’язку (КЗЗ) надходить в КП, котрий оцінює ступінь відхилення синтезується необхідний керуючий сигнал передача його до ВП по каналу прямого зв’язку (КПЗ) зміна діяльності органів-ефекторів (ВП) відновлення потрібної величини РП (усунення відхилення). За рахунок надходження до замкнутого контуру регуляції інформації по каналу зовнішнього зв’язку здійснюється регуляція “за збуреням”. Збуреня – певний зовнішній чинник, інформація про дію якого сприймається рецепторами і передається до КП. Регуляція “за збуреням” може відбуватися двома шляхами: 1. Збуреня може змінити РП відносно потрібного значення. КП, отримуючи про це сигнал по контуру зв’язку, синтезує керуючий сигнал, котрий змінює діяльність органів-ефекторів (ВП) таким чином, що потрібне значення РП відновлюється. Наприклад, при дії на організм низької температури, інформація про це сприймається холодовими терморецепторами (СП2) передається до центру терморегуляції (КП), який викликає зміну діяльності виконавчого пристрою, завдяки якому зменшуються процеси тепловіддачі та активізуються процеси теплоутворення підтримується стала температура ядра тіла, не дивлячись на дію зовнішнього чинника. 2. Збуреня може вимагати зміни РП в певному напрямку. КП, отримавши про це інформацію по КЗЗ, синтезує керуючий сигнал, котрий по КПЗ передається до ВП діяльність органів-ефекторів змінюється так, що РП змінює свою величину в потрібному напрямку. Наприклад, гемодинамічний центр (КП) при фізичній роботі (збуреня) отримує інформацію від пропріорецепторів працюючих м’язів (СП2). Ця інформація сигналізує КП про те, що у зв’язку з фізичною роботою необхідно змінити РП (системний артеріальний тиск). КП виробляє керуючий сигнал КПЗ направляє його до ВП серце скорочується сильніше і частіше – збільшення системного артеріального тиску (САТ). Інформацію про зміну САТ, про її відповідність рівню фізичного навантаження, КП отримує по КЗЗ. 2. Поняття про рефлекс. Будова рефлекторної дуги та її ланок. Нервова регуляція функцій організму – здійснюється за участю нервової системи (частіше, та не завжди – ЦНС). Особливостями нервової регуляції є те, що вона відбувається: а) точно; б) швидко; швидко вмикається, швидко вимикається, якщо потреба в ній зникає. Елементарним, найпростішим актом нервової регуляції є рефлекс – відповідь на подразнення, що починається з подразнення рецепторів і здійснюється за участю нервової системи. Рефлекторна дуга – шлях, по якому передається інформація при здійсненні рефлексу. Тобто, рефлекторна дуга – морфологічний субстрат рефлексу. Схема найпростішої, елементарної рефлекторної дуги (на прикладі шкірно-м’язового рефлексу) має такий вигляд: Із схеми видно, що рефлекторна дуга має такі відділи: 1 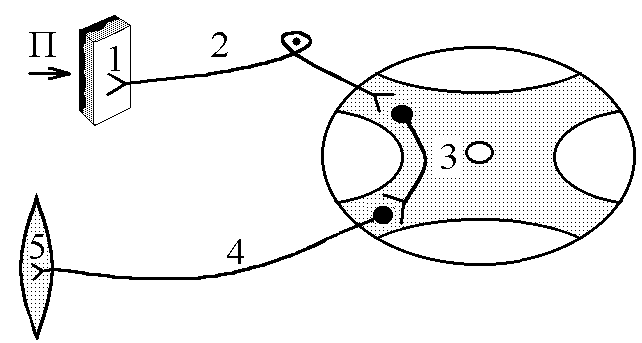 . Рецептори певної рефлексової зони. Вони забезпечують: . Рецептори певної рефлексової зони. Вони забезпечують:- сприймання інформації про дію подразника; - первинний аналіз цієї інформації (якість, сила, час дії, ступінь новизни подразника); - кодування інформації – перетворення енергії подразника в енергію нервового сигналу (серія ПД), причому ця серія ПД відображає параметри подразника. 2. Аферентний провідник – провідник першого порядку – чутливі (аферентні) нервові волокна, відростки псевдоуніполярних клітин, що локалізовані в чутливих гангліях. Забезпечують передачу інформації про дію подразника в нервовий центр: а) точно, б) швидко, в) без змін. 3. Нервовий центр – структури у межах ЦНС, що беруть участь у здійсненні рефлексу. Забезпечують аналіз інформації, що надійшла з рецепторів, і синтез адекватного еферентного сигналу – аналітико-синтетична функція. На основі цієї функції ЦНС як сукупність нервових центрів виконує координаційну функцію – узгоджує між собою діяльність різних структур, органів, тканин організму. 4. Еферентний провідник – провідник другого порядку – передає від нервового центру керуючий (еферентний) сигнал до органа-ефектора: а) швидко, б) точно, в) без змін. 5. Орган-ефектор – змінює свою діяльність під впливом керуючого сигналу так, що досягається пристосувальний (корисний) результат. Рефлекси замикаються на різних рівнях ЦНС, аж до кори головного мозку. Відповідно до цього існує поняття про інтегральний та локальний центри. Л 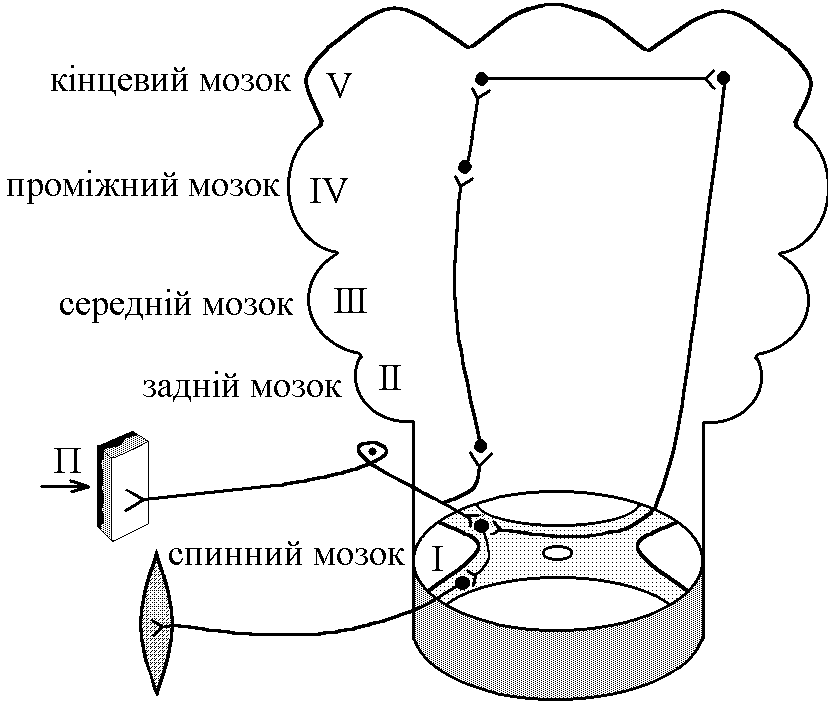 окальний нервовий центр – сукупність структур ЦНС, без яких здійснення даного рефлексу стає неможливим (якщо зруйнувати будь-яку з них, рефлекторна реакція зникає). окальний нервовий центр – сукупність структур ЦНС, без яких здійснення даного рефлексу стає неможливим (якщо зруйнувати будь-яку з них, рефлекторна реакція зникає).Інтегральний нервовий центр сукупність усіх нейронів ЦНС, що беруть участь у здійсненні даного рефлексу. Таким чином, локальний центр входить до складу інтегрального. Нейрони локального центру суворо специфічні для кожного рефлексу. Нейрони інтегрального центру можуть бути спільними для різних рефлексів. Тому ці нейрони сприяють координації (інтеграції) окремих рефлексів у цілісні реакції організму у відповідь на подразнення (узгоджують окремі рефлекси між собою). 3. Рецептори, їх класифікація та збудження. Рецептори – спеціалізовані структури, що забезпечують: а) сприйняття інформації про дію подразника; б) первинний аналіз цієї інформації (сила, якість, час дії, новизна подразника). в) кодування інформації. Існують різні класифікації рецепторів, в основі яких лежать різні властивості рецепторів: 1. За психофізіологічними відчуттями, що виникають при збудженні рецепторів, вони поділяються на:
2. За особливостями будови: 2.1. За наявністю спеціалізованої сенсорної клітини:
2.2. За наявністю чи відсутністю допоміжних структур:
3. За природою адекватного подразника:
4. За локалізацією:
5. За швидкістю адаптації:
У структурі більшості рецепторів виділяють такі елементи: - власне сприймаюча структура – нервове закінчення (первинні рецептори) чи спеціалізована сенсорна клітина (вторинні), що забезпечує сприйняття інформації про дію подразника; - генеруюча структура – найближчий до нервового закінчення перехват Ранв’є, що генерує серію ПД (аферентний сигнал) про дію подразника; 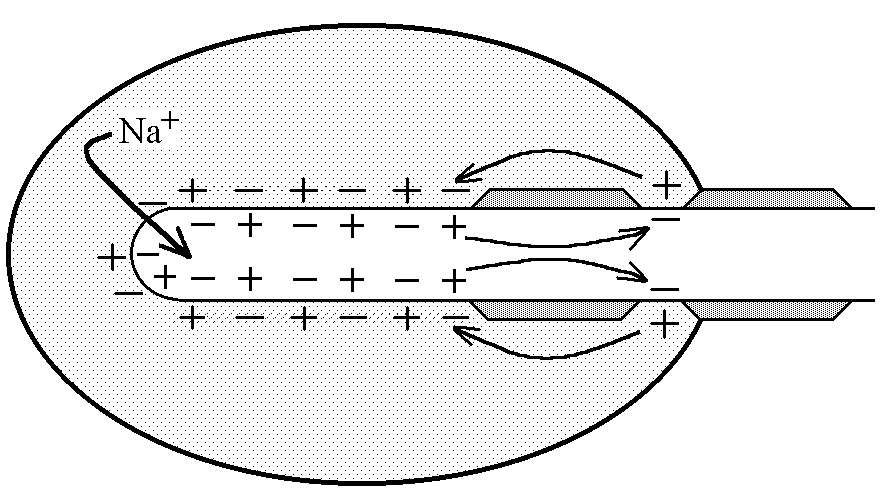 - допоміжний апарат – сукупність структур, що забезпечують найоптимальніші умови для сприйняття рецептором інформації про дію адекватного подразника. Допоміжний апарат може бути влаштований дуже просто (сполучнотканинна капсула) чи дуже складно (усі структури ока окрім паличок та колбочок). - допоміжний апарат – сукупність структур, що забезпечують найоптимальніші умови для сприйняття рецептором інформації про дію адекватного подразника. Допоміжний апарат може бути влаштований дуже просто (сполучнотканинна капсула) чи дуже складно (усі структури ока окрім паличок та колбочок).Розглянемо механізм збудження рецепторів на прикладі тільця Фатер-Пачіні. П 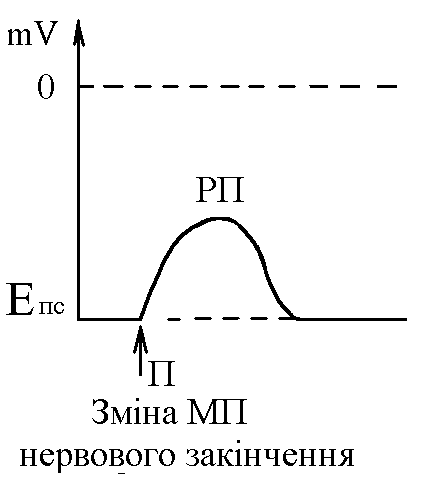 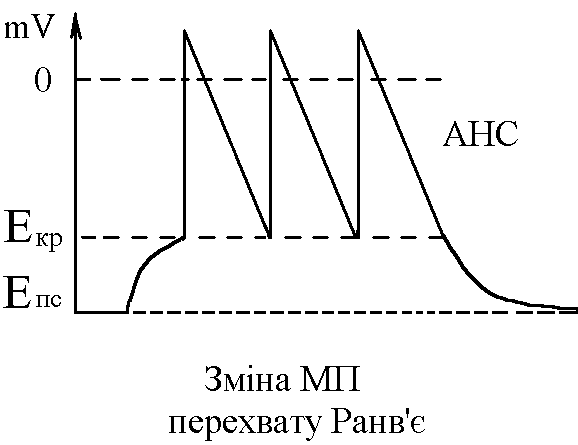 ри дії на рецептор адекватного подразника (П) – тиск на шкіру деформується його капсула (допоміжний апарат) деформація мембрани нервового закінчення (сприймаюча структура) зміна проникності мембрани для іонів натрію вхід іонів до нервового закінчення за градієнтом концентрації деполяризація мембрани місцеве збудження, яке ще називають рецепторним потенціалом (РП). Цей потенціал, як і будь-яке місцеве збудження підкоряється законам силових відношень, сумується, поширюється на малу відстань за рахунок місцевих струмів, супроводжується підвищенням збудливості мембрани. ри дії на рецептор адекватного подразника (П) – тиск на шкіру деформується його капсула (допоміжний апарат) деформація мембрани нервового закінчення (сприймаюча структура) зміна проникності мембрани для іонів натрію вхід іонів до нервового закінчення за градієнтом концентрації деполяризація мембрани місцеве збудження, яке ще називають рецепторним потенціалом (РП). Цей потенціал, як і будь-яке місцеве збудження підкоряється законам силових відношень, сумується, поширюється на малу відстань за рахунок місцевих струмів, супроводжується підвищенням збудливості мембрани.РП поширюється на сусідні ділянки мембрани за рахунок місцевих струмів, що мають напрямок від “+” до “-” (катодний) і у першому перехваті Ранв’є спричиняють деполяризацію мембрани перехвату; якщо деполяризація дійде до критичного рівня, на мембрані виникає серія ПД – аферентний нервовий сигнал. М 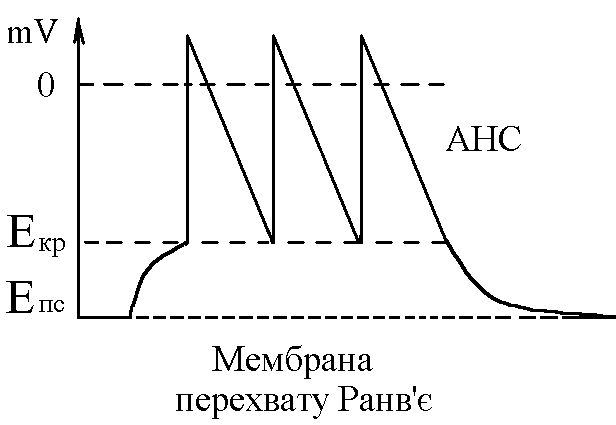 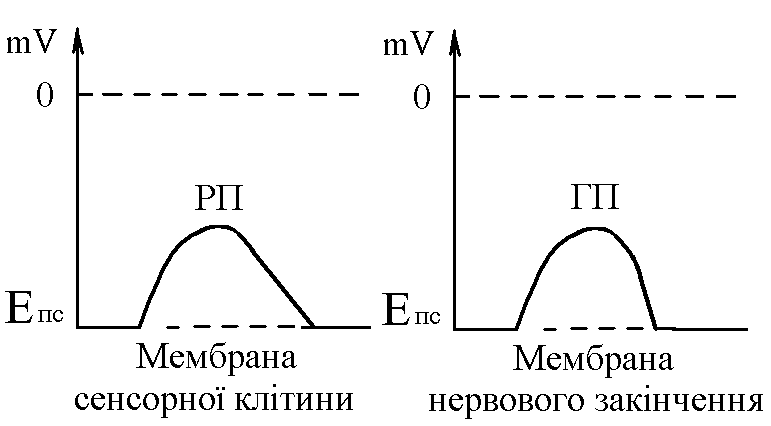 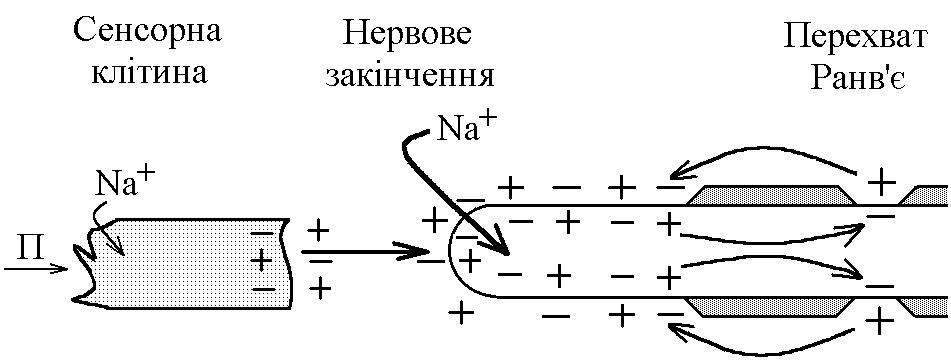 еханізм збудження вторинних рецепторів принципово не відрізняється від вищеописаного. Проте, інформація про дію подразника сприймається спеціалізованою сенсорною клітиною. Під впливом подразника мембрана клітини збільшує проникність для Na+ вхід їх до клітини деполяризація мембрани (місцеве збудження, РП) виділення клітиною медіатора в синаптичну щілину дифузія його до мембрани нервового закінчення взаємодія з мембранними циторецепторами нервового закінчення збільшення проникності мембрани для Na+ вхід їх до нервового закінчення деполяризація мембрани закінчення (місцеве збудження, генераторний потенціал, ГП). ГП за допомогою місцевих струмів поширюється на мембрану перехвату (катодний напрямок). Це деполяризує цю мембрану. Якщо деполяризація досягає критичного рівня, виникає серія ПД. При збудженні рецепторів ПД виникає лише у перехватах Ранв’є. Решта структур рецептора (мембрана нервового закінчення, мембрана сенсорної клітини) відповідають на подразнення лише місцевим збудженням (РП, ГП). Це важлива особливість збудження рецепторів, оскільки властивості місцевого рецептора забезпечують аналіз інформації вже на рівні рецепторів. еханізм збудження вторинних рецепторів принципово не відрізняється від вищеописаного. Проте, інформація про дію подразника сприймається спеціалізованою сенсорною клітиною. Під впливом подразника мембрана клітини збільшує проникність для Na+ вхід їх до клітини деполяризація мембрани (місцеве збудження, РП) виділення клітиною медіатора в синаптичну щілину дифузія його до мембрани нервового закінчення взаємодія з мембранними циторецепторами нервового закінчення збільшення проникності мембрани для Na+ вхід їх до нервового закінчення деполяризація мембрани закінчення (місцеве збудження, генераторний потенціал, ГП). ГП за допомогою місцевих струмів поширюється на мембрану перехвату (катодний напрямок). Це деполяризує цю мембрану. Якщо деполяризація досягає критичного рівня, виникає серія ПД. При збудженні рецепторів ПД виникає лише у перехватах Ранв’є. Решта структур рецептора (мембрана нервового закінчення, мембрана сенсорної клітини) відповідають на подразнення лише місцевим збудженням (РП, ГП). Це важлива особливість збудження рецепторів, оскільки властивості місцевого рецептора забезпечують аналіз інформації вже на рівні рецепторів.Механізм кодування інформації у рецепторах. Адаптація рецепторів. Кодування інформації у рецепторах – процес перетворення енергії подразника в енергію нервового сигналу (серія ПД). Причому параметри серії ПД (частота, тривалість, тощо) відображають параметри подразника (його силу, тривалість дії). Процес кодування інформації у рецепторах пов’язаний з процесом аналізу інформації у них і оснований на цьому аналізі. Аналіз інформації та кодування у рецепторах пов’язані з їх властивостями та здійснюються таким чином: 1. Про характер подразника – завдяки тому, що рецептори мають високу чутливість до адекватного подразника. Більшість рецепторів є полімодальними, тобто реагують на дію не лиши адекватного подразника, а й інших подразників. Проте чутливість їх до адекватного подразника висока, а до решти – ні. Тому при дії будь-якого подразника першими збуджуються ті рецептори, для яких цей подразник є адекватним, від них по неспецифічним провідним шляхам інформація надходить до відповідної зони кори – виникає відчуття. 2. Про силу подразника – аналіз та кодування відбувається завдяки таким особливостям рецепторів: 2.1. Рецептори однієї рефлексогенної зони мають різну збудливість (чутливість).При малій силі подразника реакцію збудження (серію ПД)генерують самі збудливі рецептори. При збільшенні сили подразника, кількість таких рецепторів збільшується – інформація про дію подразника передається більшою кількістю аферентних волокон. 2.2. Залежність амплітуди РП та частоти ПД аферентів від сили подразника.При збільшенні сили подразника зростає амплітуда РП (закон силових відносин) більш сильний катодний струм діє на мембрану перехвату Ранв’є виникає серія ПД з більшою частотою. Отже, використовується частотний принцип кодування інформації про силу подразника. 3. Про час дії подразника. Тривалість РП відповідає тривалості дії подразника, стільки ж триває аферентний сигнал. Тому зміна часу дії подразника призводить до зміни тривалості РП – змінюється тривалість АНС. 4. Рецептори здатні до адаптації. Вони адаптуються до подразників, якщо їх дія тривала. При цьому, хоча дія подразника триває, генеруюча структура (перехват Ранв’є) перестає генерувати серію ПД. Вважається, що основним механізмом адаптації є зменшення проникності мембрани перехвату для Na+. В 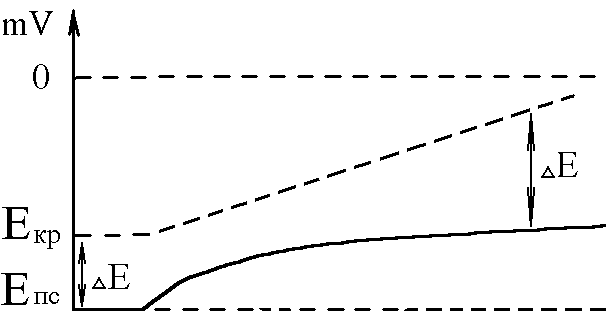 розвитку адаптації рецепторів важливе значення відіграють низхідні впливи ЦНС (центральна регуляція рівня активності рецепторів). Адаптація до тривалої дії подразника звільняє ЦНС від переробки непотрібної інформації. У рецепторах може відбувається адаптація до довготривалих подразників малої сили, які, проте, мають велике значення для організму – при цьому чутливість рецепторів до таких подразників різко зростає, наприклад підвищення чутливості зорових рецепторів в умовах темряви. В основі розвитку такої адаптації також лежать процеси, що відбуваються в самій мембрані (зміна стану каналів) рецептора. розвитку адаптації рецепторів важливе значення відіграють низхідні впливи ЦНС (центральна регуляція рівня активності рецепторів). Адаптація до тривалої дії подразника звільняє ЦНС від переробки непотрібної інформації. У рецепторах може відбувається адаптація до довготривалих подразників малої сили, які, проте, мають велике значення для організму – при цьому чутливість рецепторів до таких подразників різко зростає, наприклад підвищення чутливості зорових рецепторів в умовах темряви. В основі розвитку такої адаптації також лежать процеси, що відбуваються в самій мембрані (зміна стану каналів) рецептора.4. Пропріорецептори, їх види. Будова та функції м’язових веретен. Пропріорецептори – власні рецептори опорно-рухового апарату (м’язів, сухожилків, суглобів, зв’язок). Пропріорецептори   М’язів (м’язові веретена) Суглобових сумок Сухожилків (тільця Гольджі) Види рецепторів Адекватний подразник Деформація Розтягнення Розтягнення Ступінь та швидкість розтягнення м’язів Ступінь згинання (розгинання) в суглобі Ступінь та швидкість скорочення м’яза (так як при скороченні сухожилки розтягуються) Контролюють М 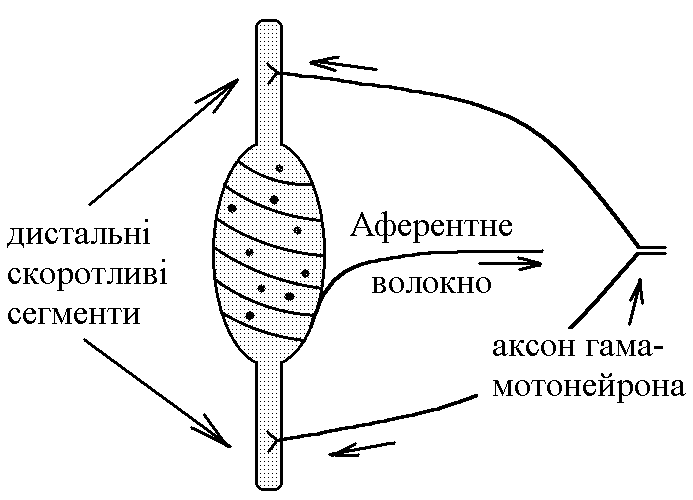 ’язові веретена – первинні механорецептори, що мають складну структуру. Кожне м’язове волокно складається з великої кількості інтрафузальних волокон (ІФВ). Ці волокна знаходяться у сполучнотканинній капсулі. Волокна є двох видів: волокно з ядерним ланцюжком та волокно з ядерною сумкою. У центрі волокна знаходяться ядра та спіралевидне нервове закінчення (власне сприймаюча структура рецептора). Окрім ядерної сумки до складу волокна входять дистальні скоротливі сегменти, що отримують інервацію від -мотонейронів спинного мозку (лежать поряд з -мотонейронами, котрі інервують екстрафузальні волокна (ЕФВ) м’яза). ’язові веретена – первинні механорецептори, що мають складну структуру. Кожне м’язове волокно складається з великої кількості інтрафузальних волокон (ІФВ). Ці волокна знаходяться у сполучнотканинній капсулі. Волокна є двох видів: волокно з ядерним ланцюжком та волокно з ядерною сумкою. У центрі волокна знаходяться ядра та спіралевидне нервове закінчення (власне сприймаюча структура рецептора). Окрім ядерної сумки до складу волокна входять дистальні скоротливі сегменти, що отримують інервацію від -мотонейронів спинного мозку (лежать поряд з -мотонейронами, котрі інервують екстрафузальні волокна (ЕФВ) м’яза). Адекватним подразником ІФВ є розтягнення центральної частини – ядерної сумки. Таке розтягнення (та збудження спіралевидного нервового закінчення, а отже, й рецептора в цілому) можливе лише в двох випадках: - 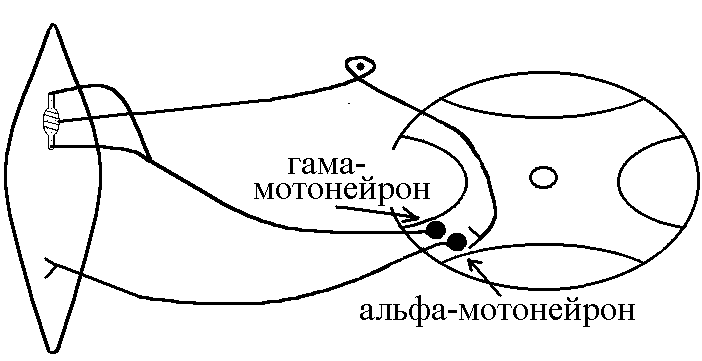 при розтягненні м’яза в цілому, так як м’язові веретена розташовані паралельно ЕФВ, а капсула прикріплена до них; при розтягненні м’яза в цілому, так як м’язові веретена розташовані паралельно ЕФВ, а капсула прикріплена до них;- без розтягнення м’яза в цілому, коли до скоротливих сегментів надходить інформація від -мотонейронів скорочення дистальних скоротливих сегментів розтягнення ядерної сумки збудження рецептора. Зауважимо, що у відповідь на збудження м’язових веретен виникають міотатичні рефлекси (рефлекси на розтягнення), при цьому міотатично активуються -мотонейрони, що інервують ЕФВ того м’яза, в якому розташоване м’язове веретено. Через -мотонейрони здійснюється центральна регуляція рівня активності м’язових веретен. Завдяки цому рецептори можуть зберігати достатній рівень збудливості (чутливості) навіть при скороченні м’яза, якщо при цьому паралельно активізуються -мотонейрони. |