1. Биология наука о жизни, об общих закономерностях существования и развития живых существ. Место и задачи предмета в системе медицинского образования. Биология
 Скачать 0.58 Mb. Скачать 0.58 Mb.
|

Регуляция экспрессии (транскрипции) генов у прокариот.Изучение регуляции генной активности упрокариот привело французских микробиологов Ф. Жакоба и Ж. Моно к созданию (1961) оперонной модели регуляции транскрипции. Оперон — это тесно связанная последовательность структурных генов, определяющих синтез группы белков, которые участвуют в одной цепи биохимических преобразований. Например, это могут быть гены, которые детерминируют синтез ферментов, участвующих в метаболизме какого-либо вещества или в синтезе какого-то компонента клетки. Оперонная модель регуляции экспрессии генов предполагает наличие единой системы регуляции у таких объединенных в один оперон структурных генов, имеющих общий промотор и оператор. Особенностью прокариот является транскрибирование мРНК со всех струтурных генов оперона в виде одного полицистронноготранскрипта, с которого в дальнейшем синтезируются отдельные пептиды. Примером участия генетических и негенетических факторов в регуляции экспрессии генов у прокариот может служить функционирование лактозного оперона у кишечной палочки Е. colt. При отсутствии в среде, на которой выращиваются бактерии, сахара лактозы активный белок-репрессор, синтезируемый геном-регулятором (I), взаимодействует с оператором (О), препятствуя соединению РНК-полимеразы с промотором (Р) и транскрипции структурных генов Z, Y, А. Появление в среде лактозы инактивирует репрессор, он не соединяется с оператором, РНК-полимераза взаимодействует с промотором и осуществляет транскрипцию полицистронноймРНК. Последняя обеспечивает синтез сразу всех ферментов, участвующих в метаболизме лактозы. Уменьшение содержания лактозы в результате ее ферментативного расщепления приводит к восстановлению способности репрессора соединяться с оператором и прекращению транскрипции генов Z, Y, А. Таким образом, регуляция экспрессии генов, организованных у прокариот в опероны, является координированной. Синтез полицистронноймРНК обеспечивает одинаковый уровень синтеза всех ферментов, участвующих в биохимическом процессе. Регуляция экспрессии генов у эукариотВ связи с особенностями организации отдельных генов эукариот и генома в целом регуляция генной активности у них характеризуется некоторыми отличиями по сравнению с прокариотами. Уэукариот не установлено оперонной организации генов. Гены, определяющие синтез ферментов одной цепи биохимических реакций, могут быть рассеяны в геноме и, очевидно, не имеют, как упрокариот, единой регулирующей системы (ген-регулятор, оператор, промотор). В связи с этим синтезируемые мРНКу эукариот моноцистронны, т.е. являются матрицами для отдельных пептидных цепей.В настоящее время механизмы регуляции и координации активности эукариотических генов интенсивно изучаются. Установлено, что их функционирование несомненно подчиняется регуляторным воздействиям, однако регуляция транскрипции у эукариот является комбинационной, т.е. активность каждого гена регулируется большим спектром генов-регуляторов У многих эукариотических генов, кодирующих белки и транскрибируемых РНК-полимеразой II, в ДНК имеется несколько областей, которые узнаются разными белками-регуляторами. Одной из них является область, расположенная вблизи промотора. Она включает около 100 пар нуклеотидов, в том числе ТАТА-блок, располагающийся на расстоянии 25 пар нуклеотидов от точки начала транскрипции. Установлено, что для успешного присоединения РНК-полимеразы II к промотору необходимо предварительное соединение с ТАТА-блоком особого белка —фактора транскрипции — с образованием стабильного транскрипционного комплекса. Именно этот комплекс ДНК с белком узнается РНК-полимеразой II. Последовательности нуклеотидов, примыкающие к ТАТА-блоку, формируют требуемый для транскрипции элемент, расположенный перед промотором. Другая область, играющая важную роль в регуляции активности эукариотических генов, располагается на большом расстоянии от промотора (до нескольких тысяч пар нуклеотидов) и называется энхансером (от англ. enhance —усиливать). И энхансер, и препромоторный элемент эукариотических генов содержат серию коротких нуклеотидных последовательностей, которые связываются с соответствующими регуляторными белками. В результате взаимодействия этих белков происходит включение или выключение генов. Особенностью регуляции экспрессии эукариотических генов является также существование белков-регуляторов, которые способны контролировать транскрипцию многих генов, кодирующих, возможно, другие белки-регуляторы. В связи с этим некоторые (главные) белки-регуляторы обладают координирующим влиянием на активность многих генов и их действие характеризуется плейотропным эффектом. Примером может служить существование белка, который активирует транскрипцию нескольких специфических генов, определяющих дифференцировку предшественников жировых клеток. Ввиду того что в геноме эукариот имеется много избыточной ДНК, а в каждой клетке организма транскрибируется всего 7—10% генов, логично предположение о том, что у них преобладает позитивный генетический контроль, при котором активация небольшой части генома оказывается более экономичной, нежели репрессия основной массы генов. Несомненной особенностью регуляции транскрипции уэукариот является подчиненность этих процессов регулирующим влияниям со стороны гормонов организма. Последние часто играют роль индукторов транскрипции. Так, некоторые стероидные гормоны обратимо связываются особыми белками-рецепторами, образуя с ними комплексы. Активированный гормоном рецептор приобретает способность соединяться со специфическими участками хроматина, ответственными за регуляцию активности генов, в которых рецепторы узнают определенные последовательности ДНК. Специфичность регулирующего воздействия гормона на транскрипцию обусловлена не только природой самого гормона, но и природой клетки-мишени, синтезирующей специфический белок-рецептор, который влияет на транскрипцию определенного для данной клетки набора генов. Примером участия гормонов в регуляции активности определенных генов может служить влияние тестостерона на развитие тканей организма по мужскому типу при наличии специфического белка-рецептора. Отсутствие последнего при мутации соответствующего гена не дает возможности гормону проникнуть в ядра клеток-мишеней и обеспечить включение определенного набора генов: развивается синдром тестикулярной феминизации, или синдром Морриса Следующая особенность регуляции генной активности уэукариот связана с образованием стойкого комплекса ДНК с белками — хроматина Ведущая роль в компактизации ДНК принадлежит гистонам, поэтому они, несомненно, участвуют и в процессах регуляции генной активности (см. разд. 3.5.4).Непременным условием для осуществления транскрипции у эукариот является предварительная декомпактизация хроматина на соответствующем участке, где временно утрачивается связь с Hi-гистонами и несколько ослабляется связь с нуклеосомными гистонами. Правда, нуклеосомная организация хроматина не утрачивается даже в ходе транскрипции, однако контакт ДНК и негистоновых белков становится возможным и происходит дерепрессия гена. Отличительной особенностью регуляции экспрессии генов уэукариот является возможность ее осуществления не только на стадии транскрипции, но и на других этапах растянутого во времени процесса реализации наследственной информации. Регуляция на стадии транскрипции является наиболее экономичной, но недостаточно быстро реагирующей на изменение ситуации. Так, возникшая в клетке потребность в каком-либо белке не может быть быстро удовлетворена путем включения транскрипции соответствующего гена. Синтезированный транскрипт должен подвергнуться процессингу, затем зрелая мРНК должна выйти из ядра в цитоплазму и, образуя комплекс с рибосомами, осуществить трансляцию информации, синтезировав пептид, который, лишь пройдя посттрансляционное изменение, формирует активный белок, необходимый клетке. В том случае, когда клетке нужно прекратить синтез какого-то продукта, после выключения транскрипции соответствующего гена в цитоплазму некоторое время будут продолжать поступать созревающие молекулы мРНК, осуществляющие там синтез пептидных цепей, пока они не деградируют под действием ферментов. Таким образом, для эффективной регуляции экспрессии генов уэукариот должны существовать механизмы, работающие не только на стадии транскрипции, но и на других этапах этого процесса. Связанная с экзон-интронной организацией генов необходимость процессичга, в том числе сплайсинга, делает возможным регуляцию этих процессов в ядре. В настоящее время обсуждается роль интронных участков ДНК в изменении схемы сплайсинга при синтезе антител (см. разд. 3.4.3.2) или цитохрома b Это создает возможность, используя один и тот же первичный транскрипт, обеспечивать образование матриц для разных пептидов, вырезая из них разные последовательности или изменяя последовательности на 5'- и 3'-концах мРНК. Очевидно, и транспорт зрелых мРНК из ядра в цитоплазму также регулируется определенным образом, так как установлено, что лишь небольшая часть РНК, транскрибируемой с генов, после сплайсинга покидает ядро. Значительное количество ее деградирует. Возможно, это является результатом процессинга, приводящего к появлению «неправильных» матриц. Существуют механизмы, обеспечивающие регуляцию процессов синтеза пептидных цепей. Они менее экономичны, но отличаются быстротой реагирования на изменения потребностей клетки в данном белке. Регуляция трансляции осуществляется на стадии инициации путем воздействия на один из факторов инициации, катализирующий присоединение к малой субъединице рибосомы тРНК, несущей метионин (формилметионин). В результате при наличии в цитоплазме мРНК трансляции на ней не происходит. Такая ситуация наблюдается, например, при отсутствии в цитоплазме гема, что ведет к выключению трансляции глобиновых цепей гемоглобина. Наконец, регуляция процесса реализации наследственной информации может осуществляться и на стадии посттрансляционных изменений. Прекращение этих процессов обусловливает задержку в формировании активных молекул белка при наличии необходимых для этого пептидных цепей. Например, для формирования активной формы белкового гормона — инсулина — из проинсулина должны вырезаться две субъединицы. Торможение этих процессов уменьшает выход конечного активного продукта. Таким образом, рассмотренный выше пример регуляции экспрессии генов демонстрирует сложнейшие взаимосвязи, которые существуют между ними в геноме. Формирование любого признака поэтому нельзя рассматривать как результат действия одной пары аллельных генов в генотипе. В любом случае регуляция экспрессии ответственного за этот признак гена осуществляется при участии других генов. 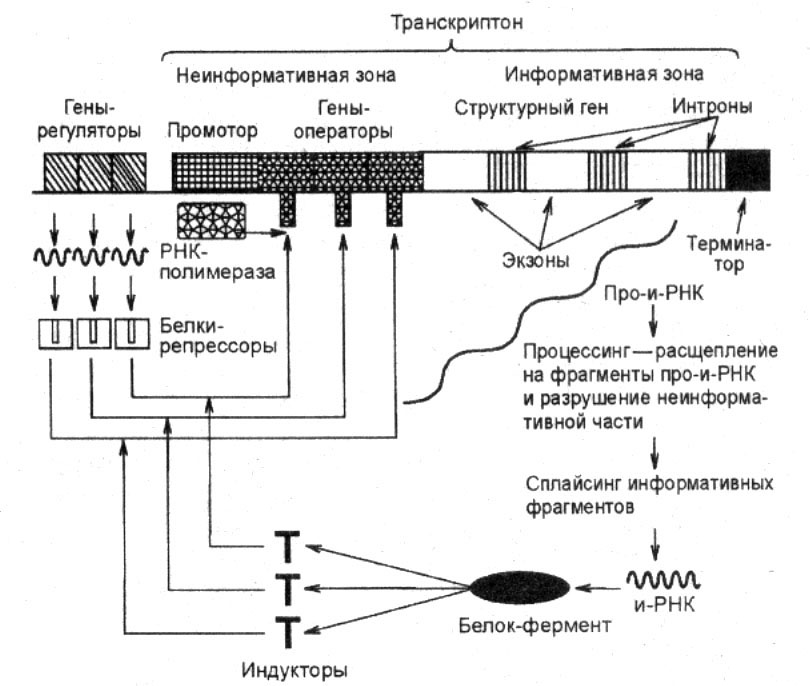 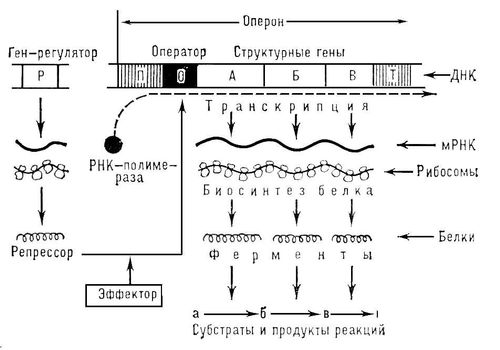 4. Кодирование и реализация наследственной информации. Этапы биосинтеза белка (транскрипция, посттранскрипционные процессы, трансляция, посттрансляционные процессы). Биосинтез белков идет в каждой живой клетке. Наследственная информация хранится в молекулах ДНК, а передается в цитоплазму информационной РНК (иРНК), которая комплементарна одной нити молекулы ДНК. Отрезок ДНК, содержащий информацию о структуре одного белка, называют геном. Свойства кода ДНК: 1. триплетность. 2. вырожденность. 3. однозначность и наличие знаков препинания. 4. неперекрываемость. 5. непрерывность. 6. универсальность. Транскрипцией называется перенос информации с ДНК на РНК. Причем матрицей для синтеза РНК служит только одна из двух цепей молекулы ДНК, называемая кодогенной цепью. Данный процесс происходит в ядре по принципу комплементарности. В транскрипции различают 4 стадии: 1. Связывание РНК-полимеразы (фермента, осуществляющего синтез РНК) с промотором. 2. Инициация – начало синтеза иРНК. В реакции участвуют нуклеотидтрифосфаты. Инициация – образование первой связи между двумя нуклеотидами (первый обычно АТФ или ГТФ). 3. Элонгация – рост цепи РНК. Для этого процесса необходимы ионы магния. В процессе элонгации образуется молекула РНК, состоящая из нуклеотидмонофосфатов, и освобождается пирофосфат (Н4Р2О7). 4. Терминация – завершение синтеза РНК в участках-терминаторах. Пройдя через поры ядерной оболочки, иРНК направляется к рибосомам, где осуществляется расшифровка генетической информации. Посттранскрипционные процессы: Внутри самого структурного гена также различают участки смысловые (экзоны) и несмысловые, «молчащие» (интроны). На матрице структурного гена синтезируется так называемая про-иРНК, копия всего гена. Затем про-иРНК подвергается созреванию, в ходе которого там же в ядре все интроны вырезаются, а концы кодирующих последовательностей соединяются. Этот процесс называется сплайсингом. Все интроны вырезаются не всегда. При изменении условий часть из них может остаться в зрелой иРНК подобно экзонам. Иногда же вырезаются какие-либо экзоны. Таким образом, один ген способен кодировать структуру нескольких белков. Трансляцией называется процесс синтеза белка на рибосомах, направляемый матрицей иРНК. Стадии: 1. Стадия активации аминокислот. Каждая аминокислота взаимодействует с молекулой АТФ под действием особого фермента, специфичного для каждой аминокислоты. Все эти ферменты носят общее название кодазы. 2. Присоединение фосфорилированных аминокислот к тРНК с образованием комплекса. Фермент при этом освобождается. 3. Собственно трансляция, или полимеризация, аминокислотных остатков с образованием пептидных связей. 4. Конформационная стадия (белок приобретает необходимую форму). Две первые стадии идут в цитоплазме. Третья стадия – в рибосомах. Посттрансляционные процессы: По окончании трансляции первая аминокислота в белковой цепи вырезается. Полученный полипептид имеет только первичную структуру(цепь), чтобы он приобрел функциональную активность, по выходу из рибосомы синтезируется вторичная(спираль), третичная(лестница), а у некоторых белков – четвертичная структура(глобула). 5. Репликация ДНК: принципы, ферментативные системы, механизмы контроля за процессом репликации ДНК. Значение репликации ДНК в ходе онтогенеза и при смене поколений. Репликация ДНК – это процесс, в результате которого из одной молекулы образуются две дочерние, полностью идентичные материнской, что обеспечивает передачу наследственной информации от поколения к поколению. Репликация осуществляется в соответствии со следующими принципами:
В процессе репликации принимает участие много белков-ферментов. ДНК-геликаза расплетает двойную спираль ДНК, разделяя ее полинуклеотидные цепи. ДНК-топоизомераза разрывает связь между остатками фосфорной кислоты и дезоксирибозы в одной из полинуклеотидных цепей ДНК, что дает ей возможность вращаться вокруг второй и снимать напряжение, вызываемое расплетением спирали и расхождением цепей в репликационной вилке. РНК-праймаза синтезирует РНК-затравки. ДНК-полимераза осуществляет непрерывный синтез лидирующей цепи и синтез фрагментов Оказаки отстающей цепи. ДНК-лигаза сшивает фрагменты после удаления РНК-затравки. для правильного синтеза существует механизмы самоконтроля: 1-механизм самокоррекции - отщепление ошибочного нуклеотида ДНК-полимеразой и замена его необходимым 2-дорепликативная репарация-вырезание поврежденного участка по принципу комплементарности( ну типа что к одной цепи не подходит то удаляется к ебням и все) 3-пострепликативная репарация - рекомбинация между двумя вновь образованными спиралями( например если между некоторыми нуклеотидами не образуются водородные связи то получается "разрывы" которые удаляются Значение в ходе онтогенеза и при смене поколения: репликация молекул ДНК в ходе онтогенеза очень важна в ходе онтогенеза происходит рост и развитие организма которые осуществляются при делении клеток путем митоза, а при смене поколений участвуют половые клетки, т.е. происходит их оплодотворение. и в том и другом случае перед делением клетка должна удвоить наследственную информацию, заложенную в молекулах ДНК этот процесс-репликация. репликация осуществляется перед каждым делением. она начинается с раскручивания спирали ДНК в S-периоде интерфазы под действием фермента ДНК-полимеразы. Организацию наследственного аппарата клеток человека можно разделить на уровни: генный, хромосомный, геномный. Единицей генного уровня является ген. Ген –участок ДНК, кодирующий синтез одной полипептидной цепи аминокислот (одной молекулы белка) размеры гена определяются числом триплетов нуклеотидов. Таким образом на генном уровне можно проследить проявление какого-то одного признака( 1 белок-1 признак) Единицей хромосомного уровня является хромосома. Хромосома-сложное образование внутри ядра, состоит из: ДНК, белков, РНК, липидов, углеводов. В одной хромосоме размещается (локализуется) огромное кол-во генов. Таким образом, на хромосомном уровне можно наблюдать развитие многих признаков( в 1 хромосоме огромное кол-во генов=> кодируется большое кол-во белков=> большое кол-во признаков определяемые этими белками). Хромосомы имеют разную форму. Форма хромосомы определяется положением центромеры (первичной перетяжки, к которой присоединяются нити веретена деления в митозе). Если центромера делит хромосому пополам, то у неё образуются равные плечи, поэтому такую хромосому называют «равноплечей» или метацентрической. Если центромера немножко смещена в сторону одного плеча – это «неравноплечая» или субметацентрическая хромосома. Если центромера делит хромосому так, что одно плечо короче другого на 75%, то её называют «резко неравноплечая» или – акроцентрическая. Если же центромера располагается в одном конце хромосомы, то хромосому называют телоцентрической. Совокупность хромосом ядра, их число, форма и структура называется кариотипом. У человека кариотип 2n=46 был установлен в 1956г. двумя учёными: Дж. Тийо и А. Леваном. В кариотипе мужчин и женщин есть одинаковые хромосомы, их большинство – 44 – это неполовые хромосомы или аутосомы (44А); и есть одна пара хромосом (23), по которой отмечается различие: у женщин ХХ, у мужчин ХY. Геном-совокупность всех генов в гаплоидном наборе, имеющихся у организмов определенного вида. Кариотип человека изображают в виде схемы, на которой хромосомы располагают в ряд по мере убывания их длинны и по одной из каждой пары. Все хромосомы объединены в 7 групп. Распределены хромосомы на схеме с учётом размеров хромосом и локализации центромерного участка, и каждая хромосома имеет свой номер (арабская цифра). Хроматин (хроматиновая нить) представляет собой интерфазное состояние хромосомы. Поэтому число хроматиновых нитей в соматических клетках должно соответствовать диплоидному набору хромосом. Хроматин - это функционально активное состояние хромосом. Хроматин, также как и хромосома, неоднороден по своей структуре. Различают два типа хроматина: эухроматин и гетерохроматин, которые морфологически и функционально отличаются друг от друга. Эухроматин - это деспирализованные и функционально активные участки хроматина, в этих участках интенсивно происходят процессы транскрипции. Гетерохроматин - более спирализованные и функционально неактивные участки хроматина. Эти участки содержат незначительное количество структурных генов и, по существу, представляют собой участки хроматина временно или постоянно выключенные из процессов регуляции клеточной активности. Эухроматин по своей структуре является конденсированным, на нем не может происходить синтез РНК, но он участвует в синтезе ДНК. Локализуется в центромере Эухроматин в свою очередь можно разделить на 2 вида: активный и факультативный гетерохроматин. Активный эухроматин по структуре диффузный на нем идет синтез как ДНК так и РНК локализуется в плечах хромосом. Факультативный находится в конденсированном состоянии, участвует в синтезе ДНК, но не участвует в синтезе РНК, локализуется в плечах хромосом.
|