экзамен магистратура экология. 1. Экология как наука. Место экологии в структуре человеческого знания. В начале xx в сформировалась новая биологическая наука экология. Термин экология
 Скачать 2.4 Mb. Скачать 2.4 Mb.
|

|
34. Концепция экологической ниши. Фундаментальная и реализованная ниши. Для определения роли, которую играет тот или иной вид в экосистеме американским зоологом – Дж. Гринеллом в 1917 году введено понятие «экологическая ниша». Гринелл термином «ниша» определял самую мелкую единицу распространения вида. Английский эколог Ч. Элтон (1927) описывал «нишу» как место данного организма в биотической среде, его положение в цепях питания. Классическое определение экологической ниши дал американский эколог Дж. Ивлин Хатчинсон. Согласно сформулированной им концепции, экологическая ниша представляет собой часть воображаемого многомерного пространства, отдельные измерения которых соответствуют факторам, необходимым для нормального существования вида. Экологическую нишу, определяемую только физиологическими особенностями организмов, Дж. Хатчинсон назвал фундаментальной, а ту, в пределах которой вид реально встречается в природе – реализованной. Допущения модели: Реакция на один фактор не зависит от воздействия другого фактора; Независимость факторов друг от друга; Пространство внутри ниши однородное с одинаковой степенью благоприятности. Это различие подчёркивает, что межвидовая конкуренция приводит к снижению плодовитости и жизнеспособности и что в фундаментальной экологической нише может быть такая часть, занимая которую вид в результате межвидовой конкуренции не в состоянии больше жить и успешно размножаться. Эта часть фундаментальной ниши вида отсутствует в его реализованной нише[7]. Таким образом, реализованная ниша всегда входит в состав фундаментальной или равна ей. Экологическая ниша – это абстрактное понятие, это совокупность всех факторов среды, в пределах которых возможно существование видов в природе. Она включает химические, физические, физиологические и биотические факторы, необходимые организму для жизни, и определяется его морфологической приспособленностью, физиологическими реакциями и поведением. По образному выражению Ю. Одума, термин «экологическая ниша» относится к роли, которую играет организм в экосистеме. Иначе говоря, местообитание – это адрес, тогда как ниша – «профессия» вида. Чтобы охарактеризовать экологическую нишу вида, необходимо знать, чем он питается и кто его поедает, способен ли он к передвижению и, наконец как он воздействует на другие элементы экосистемы (биогеоценоза) Положение вида, которое он занимает в обшей системе биоценоза, включая комплекс его биоценотических связей и требований к абиотическим факторам среды, называютэкологической нишей вида. Концепция экологической ниши оказалась очень плодотворной для понимания законов совместной жизни видов. Понятие «экологическая ниша» следует отличать от понятия «местообитание». В последнем случае подразумевается та часть пространства, которая заселена видом и которая обладает необходимыми абиотическими условиями для его существования. Экологическая ниша вида зависит не только от абиотических условий среды, но и в не меньшей мере от его биоценотического окружения. Это характеристика того образа жизни, который вид может вести в данном сообществе. Сколько на Земле видов живых организмов — столько же и экологических ниш. Перекрывание ниш происходит тогда, когда две популяции используют одни и те же ресурсы. По терминологии Хатчинсона это означает, что каждый n-мерный гиперобъем включает в себя часть другого или, другими словами, что некоторые точки двух множеств, которые составляют их реализованные ниши, идентичны. Перекрывание считается полным, если две организменные единицы характеризуются идентичными нишами; если две ниши полностью различны, то перекрывание не наблюдается. Обычно ниши перекрываются только частично, при этом одни ресурсы являются общими, а другие используются исключительно одной или другой организменной единицей. Допускается, что конкуренция интенсивна и приводит к выживанию только одного вида в оспариваемом пространстве ниши. Возможны следующие случи перекрывания ниш (рис. 3). Во-первых, одна фундаментальная ниша может целиком располагаться внутри другой и тогда исход конкуренции будет зависеть от относительной конкурентоспособности каждого вида. Если вид 2, ниша которой расположена внутри ниши вида 1, не имеет превосходства в конкуренции, то он будет вытеснен (элиминирован) из сообщества. Если же вид 2 более конкурентоспособен, то реализованная ниша вида 1 сократится, и оба вида будут сосуществовать, причем реализованная ниша доминирующего конкурента будет располагаться внутри фундаментальной ниши вида 1. 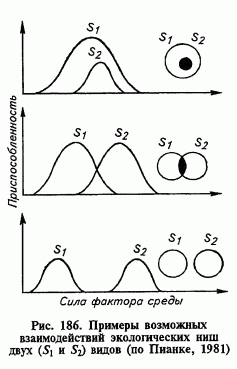 Во-вторых, две фундаментальные ниши могут перекрываться не полностью. В данном случае важным является положение зоны перекрывания к зоне оптимумов на кривой толерантности видов. Если перекрывание распространяется на зону оптимума, то сосуществование видов невозможно. Менее конкурентный вид будет элиминирован из сообщества. Во тором случае, когда перекрывание закрагивает пессимальные зоны кривых толерантности, возможно неустойчивое состояние сосуществования видов. Неустойчивость определяется изменениями окружающей среды, в случае отклонения каких-то параметров от многолетних показателей один из видов может получить конкурентное преимущество, что приведет к элиминации не конкурентного на этот момент вида. В-третьих, фундаментальные ниши могут только соприкасаться друг с другом и, хотя прямой конкуренции при этом не происходит, такая ситуация может быть следствием ее избегания. Наконец, если две фундаментальные ниши полностью разделены (перекрывание отсутствует), конкуренция также происходить не может и обе организменные единицы полностью занимают свои фундаментальные ниши. Основным недостатком изложенных рассуждении является тот факт, что в природе ниши часто перекрываются, а конкурентное исключение не происходит. Само по себе перекрывание, очевидно, еще не обусловливает конкуренции. Перекрывание используемых местообитаний может попросту свидетельствовать о том, что конкуренты изолировались друг от друга каким-то иным способом. Если же количество ресурсов имеется в избытке, две организменные единицы могут потреблять их одновременно без вреда друг для друга. На деле сильное перекрывание ниш нередко может быть связано со слабой конкуренцией подобно тому, как полное разделение ниш служит свидетельством избегания конкуренции в тех случаях, где она потенциально может быть сильной (например, в случае межвидовой территориальности). 35. Сукцессия. Определение. Основные виды сукцессий. Концепция климаксового состояния экосистемы. Относительно длительное существование биоценоза на одном месте (сосновый или еловый лес, низинное болото) изменяет биотоп (место, на котором существует биоценоз) так, что он становится малопригодным для существования одних видов, но пригодным для внедрения или развития других. В результате в данном биотопе постепенно развивается другой биоценоз, более приспособленный к новым условиям среды. Такая многократная смена одних биоценозов другими называется сукцессией. Сукцессия — это постепенная, необратимая, направленная смена одних биоценозов другими на одной и той же территории под влиянием природных факторов или воздействия человека. Термин «сукцессия» впервые употребил французский ботаник Де Люк в 1806 г. для обозначения смен растительности. Это один из ключевых терминов современной экологии. Классический пример сукцессии — зарастание озера или речной старицы и превращение ее сначала в болото, а затем, через длительный промежуток времени, в лесной биоценоз. Вначале водная гладь мелеет, затягивается со всех сторон сплавиной, на дно опускаются отмершие части растений. Постепенно зеркало воды затягивается травой. Этот процесс будет длиться несколько десятков лет, а затем на месте озера или старицы образуется верховое торфяное болото. Еще позже болото постепенно начнет зарастать древесной растительностью, скорее всего сосной. По прошествии какого-то периода времени процессы тор- фообразования на месте бывшего водоема приведут к созданию избыточного увлажнения и к гибели леса. Наконец, появится новое болото, но уже отличное от того, что было прежде. Причины сукцессий. Сукцессионные смены обычно связывают с тем, что существующая экосистема (сообщество) создает неблагоприятные условия для наполняющих ее организмов (почвоутомление, неполный круговорот веществ, самоотравление продуктами выделений или разложения и т. п.). Такие явления реальны, но не объясняют всех случаев смен экосистем. Наряду с природными факторами, причинами динамики экосистем все чаще выступает человек. К настоящему времени им разрушено большинство коренных (климаксных) экосистем. Например, степи почти полностью распаханы (сохранились только на заповедных участках). К сменам экосистем ведут также такие виды деятельности человека, как осушение болот, чрезмерные нагрузки на леса. Например, в результате отдыха населения (рекреации), химических загрязнений среды, усиленного выпаса скота, пожаров и т. п. Антропогенные воздействия часто ведут к упрощению экосистем. Такие явления обычно называют дигрессиями. Различают, например, пастбищные, рекреационные и другие дигрессии. Смены такого типа обычно завершаются не климаксными экосистемами, для которых характерно усложнение структуры, а стадиями катоценоза, которые нередко заканчиваются полным распадом экосистем. Все сукцессии можно разделить на две большие группы. Это прежде всего аллогенные сукцессии, причины которых определяются внешними влияниями (природными или антропогенными), изменяющими условия среды. Таким образом, при аллогенной сукцессии источник изменения биоценоза находится в окружающей его среде. Движущие силы сукцессии имеют направленный характер: среда → растительность. К такому типу сукцессий можно отнести переход от эвтрофного озера, обогащенного питательными веществами стоков (например, животноводческой фермы), к болоту или наземному сообществу. Сукцессии, происходящие в результате изменения условий среды самими сообществами в отсутствие постепенного изменения абиотических факторов, называются автогенными. В данном случае источником смены служит сама растительность, которая посредством изменения среды своего существования меняет структуру. Происходит как бы «самоотрицание» растительности. Схематично этот процесс можно представить следующим образом: растительность → среда → растительность. Автогенные сукцессии могут быть первичными автотрофными, вторичными автотрофными и гетеротрофными . Первичная сукцессия — это процесс развития и смены экосистем на участках, полностью свободных от растительного покрова и не сохранивших семенных зачатков растений. Образование таких участков вызывается действием ветра (ветровая эрозия или ветровая аккумуляция), отступлением берега водоема и заливанием участка водой, выгоранием растительного покрова, выгрызанием растений животными, вырубкой леса, распашкой земель, т. е. климатическими, эдафическими, биотическими и антропогенными факторами. Растения проникают на освободившуюся территорию путем переноса диаспор (семян, спор, кусочков растения) при помощи ветра, воды, животных или человека, или путем постепенного вегетативного разрастания растений, находящихся близ границ оголенной территории. В составе нового фитоценоза часто преобладают растения с диаспорами, легко разносимыми ветром, а около воды – с диаспорами, хорошо держащимися на воде. Часто, то действие, которое вызвало оголение почвы (отложение наноса водой, перевевание песка ветром) способствует и появлению на этой территории диаспор, т.е. деятельность человека. Поэтому так быстро на этих участках вырастают сорные и рудеральные растения. Заселение новой территории зависит от ряда факторов, случайных по отношению к особенностям самой территории: - от того, какие растения и как далеко они растут поблизости от нарушенного участка, - от их количества, - от направления господствующего ветра, - от высоты и силы паводка, - от качества субстрата участков, - от характера увлажнения, и т. д. Вторичная сукцессия – это восстановление экосистемы, когда-то уже существовавшей на данной территории, на местах разрушенных сообществ, где почва и часть организмов сохранились или смена одного (коренного) фитоценоза другим (производным, вторичным). Другими словами, начинается после полного или частичного нарушения экосистемы под влиянием нарушений. Если поле на месте вырубленного когда-то леса перестать обрабатывать, обычно за несколько стадий здесь вновь сформируется лесная экосистема, типичная для данного района. Процесс восстановления леса после пожара и вырубки также относится ко вторичной сукцессии. Вторичная сукцессия протекает обычно быстрее первичной, поскольку на территории, которая ранее была занята сообществом, уже имеются биогены, некоторые организмы, необходимые для обмена веществ со средой, более благоприятной для развития сообщества. Типы вторичных сукцессий (по В.М. Урусову): - антропогенная (лаборогенная) – под воздействием труда людей; - природная, или стихийная, в частности: зоогенная – вызванная животными, (например, их массовым размножением, при котором на ДВ напрочь поедаются орехи кедра корейского или съедается и вытаптывается подрост пихты цельнолистной мыши съедают его кору и луб) и фитогенная – вызванная растениями (например, завезенными чуждыми видами); - катастрофическая – вызванная природными катастрофами: пожарами (пирогенная сукцессия), ветрами, необычными паводками и т.п. Гетеротрофные сукцессии - их называют также деградационными). Эти сукцессии происходят при разложении мертвого органического вещества (детрита): трупа животного, упавшего ствола дерева, лесной подстилки и т.д. Считают, что сукцессия ведет к предельному выражению развития растительности – климаксному сообществу. Климакс – это самоподдерживающее сообщество находящиеся в равновесии с физическим местом обитания. Климакс характеризуется эдификатором. Это виды, которые в наибольшей мере создают данную среду обитания. Для климаксной системы Р/R=1. (Р – продуктивность, R – Дыхание). У концепции климакса долгая история. Один из первых исследователей сукцессии Фредерик Клементс был приверженцем теории моноклимакса и утверждал, что в любой климатической зоне существует только один истинный климакс. К его возникновению ведут все сукцессии. В конечном счете, многие экологи (в том числе Тенсли) эту теорию отвергли, и была предложена теория поликлимакса. Согласно ей климакс в данном участке может определяться одним или несколькими факторами: климатом, почвенными условиями, топографией, пожарами и т. д., поэтому в одной климатической зоне вполне может существовать целый ряд специфических типов климакса. Юджин Одум склоняется к «золотой середине» — для каждой территории характерен один единственный моноклимаксовый исход, обусловленный климатическими условиями и множество поликлимаксовыми исходами обусловленные эдафическими факторами. Классификация: Климаксное состояние сообщества разными авторами по-разному подразделяются. Так, Тенсли предлагал классифицировать их, в зависимости от сдерживающего фактора, на: *эдафические *физиографические *биотические Разумовский, который один из первых разрабатывал представления о сукцессиях предлагал делить климаксы на: *рецидивный — возникает при повторении одних и тех же нарушений, которые отбрасывают местообитание назад на одну или несколько стадий по уже пройденному пути смены *ретардационный — возникает при крайнем замедлении смены под влиянием постоянно действующих факторов не вызывающих нарушение сообщества *диаспорический — в отличие от двух предыдущих не связано с непосредственным действием внешних факторов на местообитание или сообщество. Этот тип климакса связан с отсутствием вида-эдификатора — то есть вида — строителя, чаще всего доминанта, играющего определенную роль в строительстве сообщества В 1916 Клементс создал концепцию зрелого сообщества как некой естественной единицы и даже замкнутой системы. После установления открытости сообществ, состав которых непрерывно изменяется в соответствии с градиентами условий среды, от этой концепции отказались. Место, занимаемое данным видом в серии, определяется двумя факторами: 1)скоростью, с которой вид заселяет вновь образовавшееся или разрушенное местообитание; 2)изменениями, происходящими в среде во время сукцессии. Некоторые виды медленно расселяются или медленно растут после того, как обосновались, становятся доминирующими на поздних стадиях смены ассоциаций в данной серии. Быстрорастущие растения, производящие множество мелких семян, которые разносятся на большие расстояния, обладают изначальным преимуществом над медленно расселяющимися видами и доминируют на ранних стадиях серии. Изменения, создаваемые в среде первыми колонистами, благоприятствуют вторжению в данное сообщество видов, обладающих высокой способностью к конкуренции. Сукцессия продолжается до тех пор, пока добавление к серии новых видов и исключение видов, поселившихся в данном месте ранее, уже не вызывает изменений среды развивающегося сообщества. После того как какая-либо серия дойдет до климаксной формы роста, в ее видовом составе еще происходят небольшие изменения. Идеи Клементса о том, что для каждой области характерен лишь один истинный климакс (моноклимаксная теория), вынудила ботаников создать иерархию серий, которые назывались – суб-, пре- и постклимакс: поликлимаксная теория. Уиттэкер предложил более широкую теорию мозаичного климакса – существует региональная мозаика открытых климаксных сообществ, состав которых в каждой данной местности зависит от особых условий среды в ней. Климаксное сообщество определяется многими факторами (питательные вещества, влажность, угол наклона, экспозиция и многое другое). Существуют переходящие климаксы, простейшим примером которых может служить развитие сообществ животных и растений во временных водоемах, в мертвых организмах. В простых сообществах особенности циклов развития нескольких доминирующих видов могут привести к возникновению циклического климакса. Пусть вид А может прорастать только под сенью вида В, а вид В – только под видом С, а С – только под видом А регулярный цикл доминирования видов А, С, В, А, С, В, А, С, В… - продолжительность каждой стадии будет определяться продолжительностью жизни соответствующего доминирующего вида. |