1 Физиология, ее предмет, роль задачи и формирование врачебной деятельности. Связь физиологии с другими науками. Понятие об организме, составных его элементов. Уровни морфофункциональной организации человеческого организма
 Скачать 1.07 Mb. Скачать 1.07 Mb.
|

|
26) Строение и классификация синапсов. Химический синапс. Экзоцитоз медиатора в синаптическую щель и синаптический цикл. Взаимодействие медиатора с рецепторами постсинаптической и пресинаптической мембраны. Ионатропные и метаботропные рецепторы. Ионные механизмы формирования ВПСП и ТПСП. Химические синапсы. Большинство синапсов нервной системы — химические. Функционирование таких синапсов зависит от высвобождения медиаторов. Классический химический синапс представлен пресинаптической мембраной, синаптической щелью и постсинаптической мембраной. Пресинаптическая мембрана — часть булавовидного расширения нервного окончания клетки, передающей сигнал, а постсинаптическая мембрана — часть клетки, получающей сигнал. Медиатор высвобождается из булавовидного расширения посредством экзоцитоза, проходит через синаптическую щель и связывается с рецепторами на постсинаптической мембране. Под постсинаптической мембраной расположена субсинаптическая активная зона, в которой после активации рецепторов постсинаптической мембраны происходят разнообразные биохимические процессы. В булавовидном расширении расположены содержащие медиаторы синаптические пузырьки, а также большое количество митохондрий и цистерны гладкой эндоплазматической сети. Применение традиционных методик фиксации при исследовании клеток позволяет различить на пресинаптической мембране пресинаптические уплотнения, ограничивающие активные зоны синапса, к которым при помощи микротрубочек направляются синаптические пузырьки.  Взаимодействие медиатора с рецепторами постсинаптической мембраны. • Перемещение ионов через каналы в мембране. Это приводит к изменению заряда на ней и, следовательно, мембранного потенциала. Однонаправленность передачи - нервные сигнал передаются только от пресинаптической мембраны к постсинаптической мембране. Таким образом, синапс работает по принципу клапана, что обеспечивает надежность работы нервной системы.  В возбуждающих синапсах нервной системы медиатором может являться ацетилхолин, норадреналин, дофамин, серотонин, глугаминовая кислота, вещество Р, а также большая группа других веществ, являющихся, если не медиаторами в прямом значении, то во всяком случае модуляторами (меняющими эффектиьность) синаптической передачи. Возбуждающие медиаторы вызывают появление на постсинаптической мембраневозбуждающего постсинаптического потенциала (ВПСП). Его формирование обусловлено тем, что медиатор-рецепторный комплекс активирует Na- каналы мембраны (а также вероятно и Са-каналы) и вызывает за счет поступления натрия внутрь клетки деполяризацию мембраны. Одновременно происходит и уменьшение выхода из клетки ионовК+Амплитуда одиночного ВПСП однако довольно мала, и для уменьшения заряда мембраны до критического уровня деполяризации необходима одновременная активация нескольких возбуждающих синапсов. ВПСП, образуемые на постсинаптической мембране этих синапсов, способны суммиро-ваться, т.е. усиливать друг друга, приводя к росту амплитуды ВПСП (пространственная суммация). Растет амплитуда ВПСП и при увеличении частоты поступающих к синапсу нервных импульсов (временная суммация), что повышает число выводимых в синаптическую щель квантов медиатора. Тормозной постсинаптический потенциал (ТПСП) В тормозных синапсах обычно действуют другие, тормозные, медиаторы. Среди них хорошо изученными являются аминокислота глицин (тормозные синапсы спинного мозга), гамма-аминомасляная кислота (ГАМК) — тормозной медиатор в нейронах головного мозга. Вместе с тем, тормозной синапс может иметь тот же медиатор, что и возбуждающий, но иную природу рецепторов постсинаптической мембраны. Так, для ацетилхолина, биогенных аминов и аминокислот на постсинаптической мембране разных синапсов могут существовать как минимум два типа рецепторов, и, следовательно, разные медиатор-рецепторные комплексы способны вызывать различную реакцию хемочувствительныхрецепторуправляемых каналов. Для тор-мозного эффекта такой реакцией может являться активация калиевых каналов, что вызывает увеличение выхода ионов калия наружу и гиперполяризацию мембраны. Аналогичный эффект во многих тормозных синапсах имеет активация каналов для хлора, увеличивающая его транспорт внутрь клетки. Возникающий при гиперполяризации сдвиг мембранного потенциала получил название тормозного постсинаптического. 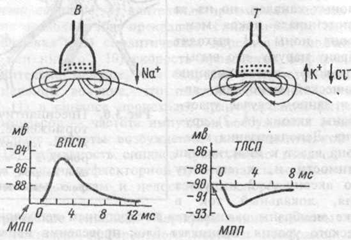 27) Аксошипиковые синапсы, их функциональная роль. Свойства химических синапсов (одностороннее проведение, синаптическая задержка и др.). Электрические синапсы, механизмы передачи возбуждения. Структурно-функциональное развитие нейронов в грудном и детском возрастах. Рост объема серого вещества мозга у детей. Свойства химических синапсов 1. Односторонняя проводимость — одно из важнейших свойств химического синапса. Асимметрия — морфологическая и функциональная — является предпосылкой для суще-ствования односторонней проводимости. 2. Наличие синаптической задержки: для того, чтобы в ответ на генерацию ПД в области пресинапса выделился медиатор и произошло изменение постсинаптического потенциала (ВПСП или ТПСП), требуется определенное время (синаптическая задержка). В среднем оно равно 0,2—0,5 мс. Это очень короткий промежуток времени, но когда речь идет о ре-флекторных дугах (нейронных сетях), состоящих из множества нейронов и синаптических связей, это латентное время суммируется и превращается в ощутимую величину — 300— 500 мс. В ситуациях, встречающихся на автомобильных дорогах, это время оборачивается трагедией для водителя или пешехода. 3. Благодаря синаптическому процессу нервная клетка, управляющая данным постси-наптическим элементом (эффектором), может оказывать возбуждающее воздействие или, наоборот, тормозное (это определяется конкретным синапсом). 4. В синапсах существует явление отрицательной обратной связи — антидромный эффект. Речь идет о том, что выделяемый в синаптическую щель медиатор может регулировать выделение следующей порции медиатора из этого же пресинаптического элемента путем воздействия на специфические рецепторы пресинаптической мембраны. Так, известно, что в адренергичсских синапсах имеются альфа2-адренорецепторы, взаимодействие с кото-рыми (норадреналин связывается с ними) приводит к снижению выделения порции норадреналина при поступлении очередного сигнала к синапсу. На пресинаптической мембране обнаруживаются рецепторы и к другим веществам. +5. Эффективность передачи в синапсе зависит от интервала следования сигналов через синапс. Если этот интервал до некоторых пор уменьшать (учащать подачу импульса по аксону), то на каждый последующий ПД ответ постсинаптической мембраны (величина ВПСП или ТПСП) будет возрастать (до некоторого предела). Это явление облегчает передачу в синапсе, усиливает ответ постсинаптического элемента (объекта управления) на очередной раздражитель; оно получило название «облегчение» или «потенциация». В основе его лежит накопление кальция внутри пресинапса. Если частота следования сигнала через синапс очень большая, то из-за того, что медиатор не успевает разрушиться или удалиться в синаптической щели, возникает стойкая деполяризация или католическая депрессия — снижение эффективности синаптической передачи. Это явление называется депрессией. Если через синапс проходит много импульсов, то в конечном итоге постсинаптическая мембрана может уменьшить ответ на выделение очередной порции медиатора. Это называется явлением десенситизации — утратой чувствительности. В определенной мере десенситизация - ответ на процесс рефрактерности (утрата возбудимости). Синапсы подвержены процессу старения. Возможно, что в основе утомления (временного падения функциональных возможностей синапса) лежат: а) истощение запасов медиатора, б) затруднение выделения медиатора, в) явление десенситизации. Т. о., утомление — это интегральный показатель. Интегративная функция нейрона. Общее изменение мембранного потенциала нейрона является результатом сложного взаимодействия (интеграции) местных ВозбПСП и ТормПСП всех многочисленных активированных синапсов на теле и дендритах клетки. На мембране нейрона происходит процесс алгебраического суммирования положительных и отрицательных колебаний потенциала. При одновременной активации нескольких возбуждающих синапсов общий ВПСП нейрона представляет собой сумму отдельных местных ВПСП каждого синапса. При одновременном возникновении двух различных синаптических влияний — ВПСП и ТПСП — происходит взаимное вычитание их эффектов. В конечном итоге реакция нервной клетки определяется суммой всех синаптических влияний. Преобладание тормозных синаптических воздействий приводит к гиперполяризации мембраны и торможению деятельности клетки. При сдвиге мембранного потенциала в сторону деполяризации повышается возбудимость клетки. Ответный разряд нейрона возникает лишь тогда, когда изменения мембранного потенциала достигают порогового значения — критического уровня деполяризации. Механизм передачи возбуждения в электрическом синапсе. Во время развития ПД происходит реверсия заряда пресинаптической мембраны. Электрический ток, возникающий между пресинаптической и постсинаптической мембраной, раздражает постсинаптическую мембрану и вызывает генерацию в ней ПД. С рождения нейроны мозга существуют по большей части независимо друг от друга. Задача мозга в течение первых 3 лет — установить и укрепить связи между ними. В это время клетки детского мозга создают по 2 миллиона новых связей — синапсов — в секунду! С развитием ребенка синапсы становятся более сложными: они растут, подобно дереву с большим количеством ветвей и веточек. Известный факт: как правило, самые первые наши воспоминания относятся к возрасту трёх и более лет. Это объясняется недостатком нейронных связей в раннем детстве. И хотя более ранние переживания «забываются», нельзя сказать, что они проходят бесследно. Ведь именно они помогают подготовить мозг к приобретению новых знаний. Когда в результате созревания мозга образуется множество нейронных связей, опыт сохраняют только активно используемые связи. К моменту половой зрелости наблюдается массовое исчезновение неиспользуемых связей. Именно здесь, на стыке задатков и воспитания, в окружающей ребёнка среде активизируются и сохраняются нервные связи. Однако если эта среда однообразная и обеднённая, нервные связи не будут использоваться и исчезнут. Это – биологическая основа раннего детского образования. Именно поэтому в раннем детстве, когда ещё востребованы излишние нервные связи, дети могут легче всего овладеть грамматикой и произношением иностранного языка. Если у человека до юношеского возраста не было никаких языковых контактов (письменных или знаковых), то он никогда не выучит ни одного слова. Развитие мозга ребёнка в первые годы жизни Для оптимального развития мозга ранние годы являются критическим (сензитивным) периодом. На протяжении всей жизни наши нервные ткани изменяются. Генетическая программа диктует нам общее строение мозга, а жизненный опыт руководит деталями. Все компоненты нейронных ансамблей новорождённого характеризуются структурной незрелостью. В первый год с максимальной скоростью происходит типизация и увеличение размеров нейронов, рост коры головного мозга. Однако в разных зонах мозга в зависимости от функциональной значимости и нагрузки можно наблюдать свой собственный темп развития клеток, клеточных ансамблей и волокнистых структур. Первыми созревают и к трём годам замедляют свой рост проекционные области коры головного мозга: двигательные, чувствительные, зрительная, слуховая, обонятельная. До двух лет наиболее активно (гораздо активней, чем у взрослого) происходит образование синапсов – мест соединения нейронов, и к семи годам их количество достигает уровня взрослого человека. Этот избыточный рост и образование синапсов имеет огромное функциональное значение, закреплённое эволюционно. Он является основой для накопления и усвоения опыта. Следовательно, мозг ребёнка функционально готов к большей нагрузке, чем это обеспечивает процесс воспитания. 28) Трофическая функция нейрона – аксонный транспорт (быстрый и медленный, антероградный и ретроградный), импульсное нейротрофическое влияние. Регенерация нервных волокон (аксонов) в ЦНС и периферической нервной системе. Структурно-функциональное изменение нейронов и глиальных клеток при старении организма. Структурно-функциональные изменения нейронов и глиальных клеток в различные периоды детства. Трофическая функция нейрона реализуется как в его отростках, так и в иннервируемых клетках. Она может осуществляться как вне прямой связи с проведением и передачей ПД (неимпульсное трофическое действие - в результате аксонного транспорта), так и непосредственно при передаче возбуждения и торможения через синапсы (импульсное трофическое явление). Аксонный транспорт осуществляет передвижение вдоль аксона нейроплазмы, органелл, макромолекул и низкомолекулярных веществ. Необходимость аксонного транспорта обусловлена тем, что по механизму диффузии тело нейрона (его трофический центр) не может обеспечить трофическое влияние на протяжении аксона, длина которого у человека может превышать 1м (например, для диффузии белка на это расстояние потребовалось бы 50 лет). Выделяют быстрый и медленный аксонный транспорт. Быстрый аксонный транспорт происходит в двух направлениях. 1)Антероградный транспорт (от тела клетки к аксонным окончаниям, V = 250 - 400 мм/сутки) доставляет везикулы, митохондрии, гликопротеины, ферменты, липиды, медиаторы и другие вещества. С промежуточной скоростью (4 — 50 мм/сутки) транспортируются митохондриальные и миозиноподобные белки, актин, клатрин, кальмодулин. 2)Ретроградный транспорт (от окончаний аксона к телу нейронов, V = 200 - 300 мм/сутки) доставляет везикулы, содержащие остатки разрушенных структур, фрагменты мембран, нейроростовые факторы, лизосомальные ферменты, ацетилхолинэстеразу, вирусы полимиелита, бешенства, столбнячный экзотоксин. Механизм быстрого транспорта: транспорт осуществляется вдоль микротрубочек с помощью белка кинозина, связывающего органеллу с микротрубочкой и преобразующего энергию АТФ в механическую работу. Медленный аксонный транспорт происходит в антероградном направлении и состоит из передвижения всего столба аксоплазмы (V = 1 - 2 мм/сутки). Этим транспортом перемещаются образованные в ЭПС белки микротрубочек и микрофиламентов, каналов, насосов, ферменты цитозоля и другие. Он определяет скорость регенерации нерва. Функциональная роль аксонного транспорта заключается в поддержании структуры и функции аксона и его окончаний, осуществлении аксонного роста и образовании новых синапсов, трофическом влиянии на иннервируемую клетку:регуляции ее обмена веществ, размножения, дифференцировки, обеспечивает тонические связи между нейронами. Импульсное нейротрофическое влияние. Образование, распространение и передача нервного импульса (ПД) изменяет не только электрические свойства клеточной мембраны, но и свойства клетки в целом. Это обусловлено тем, что в реализации биоэлектрических процессов участвуют биоэнергетические, синтетические, пластические, ультраструктурные, генетические процессы. Связывающим звеном при этом являются вторые посредники (цАМФ, ИФз, ДАГ, NO, Са²+), образующиеся при передаче в синапсах как с ионотропными, так и особенно, с метаботропными рецепторами. Примером нарушения импульсного трофического влияния, возникающего при гиподинамии организма (аксонный транспорт сохранен!), является гипотрофия скелетных мышщ (дефицит влияния через ионотропные N-холинорецепторы в синапсе) и сердца (дефицит влияния через метаботропные бета-адренорецепторы). В здоровом организме импульсное трофическое влияние и аксонный транспорт взаимодействуют друг с другом. Регенерация аксонов. Регенерация нервных волокон в ЦНС, как правило, не происходит (за исключением аксонов нейросекреторных нейронов гипоталамуса) или происходит медленно. Причины этого связаны с тем, что волокна в ЦНС не содержат базальной мембраны, необходимой для их регенерации; кроме того, олигодендроциты и астроциты выделяют факторы, тормозящие рост аксона. Гибель нейронов в ЦНС практически не компенсируется их пролиферацией (реальные возможности клеток-предшественников нейронов, обнаруженные в некоторых областях головного мозга пока мало изучены). Повреждение тела нейрона как в ЦНС, так и в ганглиях приводит к гибели и его аксона. Регенерация нервных волокон в периферической нервной системе может осушествляться, проходя предварительно этап валлеровской дегенерации. Валлеровская дегенерация. При повреждении аксона в периферической нервной системе его дистальный от места повреждения отросток погибает с образованием фрагментов аксона и миелина, которые удаляются поступившими в эту зону макрофагами (примерно в течение недели). Регенерация аксона. Базальная мембрана нервного волокна в зоне дегенерации выделает бенки (ламинин, фибронектин}, которые индуцируют рост аксона Размножающиеся леммоциты внутри цилиндра, образованного базальной мембраной волокна, формирую тэндоневральную трубку. Леммоциты образуют различные, нейротрофические факторы (фактор роста нервов, нейротрофины, глиальный нейротрофический фактор), которые стимулируют рост аксона. Нейроростовые факторы действуют преимущественно на специальные белки-репепторы клетки, которые входят в тирозинкиназную систему передачи сигнала внутрь клетки. Проксимальный отросток аксона в области травмы образует многочисленные выросты (конус роста). Те отростки, которые попадают в эндоневральную трубку, растут со скоростью 1-3 мм в сутки и образуют нервные окончания на иннервируемых клетках, остальные дегенерируют. Структурные изменения нейронов при старении организма. Происходят снижение преимущественно в коре общего числа нейронов (до 40-70%), усиливаются дистрофические процессы в нейронах и их отростках: вакуолизация, накопление липидов и пигмента липофусцина в цитоплазме, сегментарная демиелинизация аксонов, уменьшение числа аксодендритических синапсов и резервных аксошипиковых синапсов. Функциональные изменения нейронов. Снижение синтеза АТФ и активности К+, Na+ —насоса и лабильности нейронов, синтеза белка, аксонного транспорта в нейронах с накоплением «пигмента старости» липофусцина. Ослабление синтеза медиатора в синапсах, замедление проведения возбуждения через синапс, снижение функционального резерва нейронов и их трофического влияния. Структурно-функциональные изменения глии при старении организма. Относительное увеличение количества глиальных клеток по отношению к нейронам. Усиление белкового и энергетического обменов в глиоцитах, передачи пластических веществ от них к нейрону; участие глии в удалении пигмента липофусцина из нейронов. Увеличение обратного захвата глиоцитами медиаторов нейронов. Увеличение роли глиоцитов в образовании и закреплении временных связей. |