1. Иммунитет. Определение, виды и их сравнительная характеристика. Иммунитет
 Скачать 0.69 Mb. Скачать 0.69 Mb.
|

|
Простагландины Поскольку фосфолипиды всех клеточных мембран содержат арахидоновую кислоту, то и все клетки организма потенциально могут быть источником простагландинов, и они могут проявлять активность в самых разных системах. Действие ПГ проявляется чаще в тех гормоночувствительных клетках, где вторым посредником служит цАМФ: влияют на уровень цАМФ путем изменения активности аденилатциклазы. Следует подчеркнуть многофункциональность биологических эффектов ПГ - ов, причём простагландины групп Е и Г оказывают противоположное действие. Основные эффекты простагландинов
Простациклин и тромбоксан обладают прямо противоположным действием на два принципиально важных процесса: простациклин расслабляет гладкую мускулатуру сосудистой стенки и тормозит агрегацию тромбоцитов; тромбоксан, наоборот, сокращает гладкую мускулатуру сосудов и способствует агрегации тромбоцитов. Одним из основных достижений в изучении простагландинов является установление факта, что они - важнейший компонент развития воспалительного процесса. Лейкотриены Эйкозаноиды, биосинтез которых идет по липоксигеназному пути из арахидоновой кислоты. Три лейкотриена: ЛТС4, ЛТД4 и ЛТЕ4 в совокупности являются МРВ-А ( медленно реагирующим веществом, А-анафилаксии). Они секретируются тучными клетками, принимающими участие в реакциях гиперчувствительности. Эффект компонентов МРВ-А в 4000 раз сильнее, чем гистамина в отношении стимуляции гладкой мускулатуры трахеи и бронхов. Эти компоненты действуют и на гладкую мускулатуру ЖКТ, но не столь сильно. Они также действуют и на гладкую мускулатуру артерий. Как и гистамин, компоненты МРВ-А увеличивают проницаемость сосудов.[13] С -реактивный белок (СРБ) Очень чувствительный элемент крови, быстрее других реагирующий на повреждение тканей. Наличие реактивного белка в сыворотке крови - признак воспалительного процесса, травмы, проникновения в организм чужеродных микроорганизмов - бактерий, паразитов, грибов. С - реактивный белок стимулирует защитные реакции, активирует иммунитет. Содержание СРБ в сыворотке крови - до 0,5 мг/л считается нормой. Уже через 4-6 часов после того, как в организм проникает инфекция, развивается воспалительный процесс, который может сопровождаться опухолями, уровень СРБ начинает быстро расти. Чем острее воспалительный процесс, активнее заболевание, тем выше СРБ в сыворотке крови. Когда заболевание в хронической форме переходит в фазу, то содержание СРБ в крови практически не обнаруживается. Как наступает обострение, СРБ снова начинает расти. Пептиды малой плотности Пептиды малой плотности (ПМП) - относятся к новому классу регуляторных молекул, являющиеся катионными белками, открытыми в середине 80-х годов R.Lehrer и В.Н. Кокряковым. ПМП обладают широким спектром прямой противомикробной активацией, в частности подавляют рост грамположительных и грамотрицательных бактерий, грибов, некоторых вирусов. В настоящее время известны два больших класса ПМП человека: дефензины и кателецидины. Они действуют как эндогенные антибиотики и участвуют в передаче сигналов клеткам, вовлечения в процесс иммунной защиты. В основном эти белки синтезируются эпителиальными клетками покровных тканей и активированы. Недавно получены данные о том, что кателицидин LL-37 обладает прямым действием на вирус осповакцины.[7,13] Антимикробные пептиды – Лизоцим. Содержится в сыворотке, крови, слюне, слезах и других тканевых жидкостей человека. По биологической природе - это полипептид, по функциям фермент. Лизоцим действует энзиматически на клеточную стенку бактерий, расщепляя находящиеся в её составе пептидогликана и мурамовой кислоту.[4,11] Пропердин Пропердин или фактор Р-белок, содержащийся в сыворотке крови. Система пропердина состоит из самого фактора Р и 3-х дополнительных белков (А,В,0). Все они принимают участие в активации комплемента - в расщеплении С3 компонента, который в свою очередь обладает выраженными антимикробными свойствами, стимулятором фагоцитоза. Все эти субстанции имеются в организме и не нужно времени на их образование.[4] Цитокины Под термином “цитокины” объединяются так называемые ростовые факторы, которые регулируют пролиферацию, дифференцировку и функцию клеток крови, в том числе и клеток иммунной системы. Это обширный класс биохимических веществ, продуцируемый большинством свободных клеток крови, для общения друг с другом, через поверхностные рецепторы на их мембранах. Цитокины оказывают аутокринное и паракринное воздействие. 6. Цитокиновая сеть. Классификация и функция цитокинов. Цитокины - группа растворимых клеточных пептидных медиаторов, продуцирующихся разными клетками организма и играющих важную роль в обеспечении физиологических процессов в норме и при патологии. Свойства цитокинов:
Классификация цитокинов:
Классификация цитокинов по биологической активности:
7. Эндоцитозные, сигнальные и растворимые рецепторы врожденного иммунитета. Особую роль в реакциях врожденного иммунитета играют паттернраспознающие рецепторы (PRR, особенно Toll-подобные рецепторы — TLR), распознающие компоненты микроорганизмов и эндогенные сигналы опасности, которые возникают в организме. В результате действия высокоэффективных механизмов врожденная иммунная система определяет потенциальные патогены, распознавая ЛПС, пептидогликаны, липопептиды, флагеллин и многие другие консервативные и неизменяющиеся структурные молекулы. В этом отношении врожденную иммунную систему рассматривают как первую линию защиты против патогенных микроорганизмов у млекопитающих. Одна из целей врожденного иммунитета сводится к раннему установлению различий между патогенами и непатогенами, что особенно важно в пограничных тканях (слизистые оболочки пищеварительного тракта и дыхательных путей, кожа и т.д Рецепторы опознавания паттерна классифицируют по специфичности к лиганду, функции, локализации и по происхождению в эволюции. По функции они делятся на два класса: сигнальные и эндоцитозные. Сигнальные рецепторы опознавания паттерна включают, например, толл-подобные рецепторы. Эндоцитозные рецепторы опознавания паттерна, например, маннозные рецепторы макрофагов, необходимы для прикрепления, поглощения и процессирования микроорганизмов фагоцитами независимо от внутриклеточной передачи регуляторного сигнала. Кроме патогенов они опознают также апоптозные клетки. Мембранные рецепторы опознавания паттерна Рецепторы-киназы Впервые рецепторы опознавания паттерна были открыты у растений[1]. Позже множество гомологичных рецепторов было обнаружено при анализе геномов растений (у риса 370, у Arabidopsis — 47). В отличие от рецепторов опознавания паттерна у животных, которые связывают внутриклеточные протеинкиназы с помощью адапторных белков, растительные рецепторы представляют собой один белок, состоящий из нескольких доменов, внеклеточного, опознающего патоген, внутриклеточного, обладающего киназной активностью, и трансмембранного, связывающего первые два. Толл-подобные рецепторы Этот класс рецепторов опознает патогены вне клеток или в эндосомах[2]. Они были впервые обнаружены у дрозофилы и индуцируют синтез и секрецию цитокинов, необходимых для активации иммунного ответа. В настоящее время толл-подобные рецепторы обнаружены у многих видов. У животных их насчитывают 11 (TLR1-TLR11). Взаимодействие толл-подобных рецепторов с лигандами приводит к индукции сигнальных путей NF-kB и МАР-киназы, которые, в свою очередь, индуцируют синтез и секрецию цитокинов и молекул, стимулирующих презентацию антигена[3]. Цитоплазматические рецепторы опознавания паттерна Nod-подобные рецепторы Nod-подобные рецепторы — это цитоплазматические белки с различными функциями. У млекопитающих их найдено около 20, и большинство из них подразделяют на два главных подсемейства: NOD и NALP. Кроме того, к этому семейству рецепторов относят трансактиватор главного комплекса гистосовместимости класса II и некоторые другие молекулы. Опознавая патоген внутри клетки, рецепторы олигомеризуются и образуют инфламмасому, активирующую ферменты протеолитической активации цитокинов, например, интерлейкина 1 бета. Рецепторы активируют также сигнальный путь NF-kB и синтез цитокинов[4][5]. NODS Известны два главных представителя: NOD1 и NOD2. Связывают два разных бактериальных пептидогликана[6]. NALPS Известно 14 белков (NALP1 — NALP14), которые активируются бактериальными пептидогликанами, ДНК, двухцепочечной РНК, парамиксовирусом и мочевой кислотой. Мутации некоторых из NALPS являются причиной наследственных аутоиммунных заболеваний. Другие Nod-подобные рецепторы Такие молекулы, как IPAF и NAIP5/Birc1e также индуцируют протеолитическую активацию цитокинов в ответ на появление сальмонеллы и легионеллы. РНК хеликазы Индуцируют антивирусный иммунный ответ после активации вирусной РНК. У млекопитающих это три молекулы: RIG-I, MDA5 и LGP2. Секретируемые рецепторы опознавания паттерна Многие рецепторы опознавания паттерна, например, рецепторы комплемента, коллектины и пентраксины, к которым, в частности, относится C-реактивный белок, не остаются в синтезирующей их клетке и попадают в сыворотку крови[7]. Одним из важнейших коллектинов является лектин, связывающий маннозу; он опознает широкий спектр патогенов, в состав клеточной стенки которых входит манноза, и индуцирует лектиновый путь активации системы комплемента[8]. 8. Секреторные рецепторы врожденного иммунитета. Растворимые рецепторы для патогенов. Это белки крови, непосредственно связывающиеся с различными консервативными, повторяющимися углеводными или липидными структурами микробной клетки (pattern-структурами). Эти белки обладают опсоническими свойствами, некоторые из них активируют комплемент. Основную часть растворимых рецепторов составляют белки острой фазы. Концентрация этих белков в крови быстро нарастает в ответ на развитие воспаления при инфекции или повреждении тканей. К белкам острой фазы относятся: • С-реактивный белок (он составляет основную массу белков острой фазы), получивший название вследствие способности связываться с фосфорилхолином (С-полисахаридом) пневмококков. Образование комплекса С-реактивный белок- фосфорилхолин способствует фагоцитозу бактерий, поскольку комплекс связывается с Clg и активирует классический путь комплемента. Белок синтезируется в печени, и его концентрация быстро нарастает в ответ на интерлейкин-б; Липополисахарид-связывающий белок (ЛСБ, англ. lipopolysaccharide binding protein, LBP) — секретируемый белок, компонент острой фазы воспаления, который связывает с высокой афинностью бактериальный липополисахарид и усиливает связывание с ним CD14. Таким образом, ЛСБ обеспечивает первый этап в процессе моноцитарного иммунного ответа.
• сывороточный амилоид Р близок по структуре и функции к С-реактивному белку; • маннозосвязывающий лектин активирует комплемент по лектиновому пути, является одним из представителей сывороточных белков-коллектинов, распознающих углеводные остатки и действующих как опсонины. Синтезируется в печени; • белки сурфактанта легких также принадлежат к семейству коллектинов. Обладают опсоническим свойством, особенно в отношении одноклеточного гриба Pneumocystis carinii; • другую группу белков острой фазы составляют белки, связывающие железо, - трансферрин, гаптоглобин, гемопексин. Такие белки препятствуют размножению бактерий, нуждающихся в этом элементе. 9. Система комплемента Комплемент - система сывороточных белков и нескольких белков клеточных мембран, выполняющих 3 важные функции: опсонизацию микроорганизмов для дальнейшего их фагоцитоза, инициацию сосудистых реакций воспаления и перфорацию мембран бактериальных и других клеток. Компоненты комплемента обозначают буквами латинского алфавита C, B и D с добавлением арабской цифры (номер компонента) и дополнительных строчных букв. Компоненты классического пути обозначают латинской буквой «С» и арабскими цифрами (C1, C2 ... C9), для субкомпонентов комплемента и продуктов расщепления к соответствующему обозначению добавляют строчные латинские буквы (C1q, C3b и т.д.). Активированные компоненты выделяют чертой над литерой, инактивированные компоненты - буквой «i» (например, iC3b). • Активация комплемента В норме, когда внутренняя среда организма «стерильна» и патологического распада собственных тканей не происходит, уровень активности системы комплемента невысок. При появлении во внутренней среде микробных продуктов происходит активация системы комплемента. Она может происходить по трём путям: альтернативному, классическому и лектиновому. - Альтернативный путь активации. Его инициируют непосредственно поверхностные молекулы клеток микроорганизмов [факторы альтернативного пути имеют буквенное обозначение: P (пропердин), B и D]. 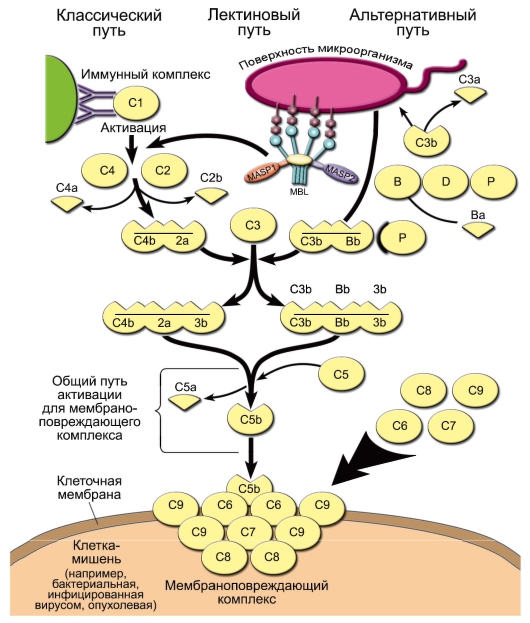 Из всех белков системы комплемента в сыворотке крови больше всего C3 - его концентрация в норме составляет 1,2 мг/мл. При этом всегда имеется небольшой, но значимый уровень спонтанного расщепления C3 с образованием C3a и C3b. Компонент C3b - опсонин, т.е. он способен ковалентно связываться как с поверхностными молекулами микроорганизмов, так и с рецепторами на фагоцитах. Кроме того, «осев» на поверхности клеток, C3b связывает фактор В. Тот, в свою очередь, становится субстратом для сывороточной сериновой протеазы - фактора D, который расщепляет его на фрагменты Ва и Bb. C3b и Bb образуют на поверхности микроорганизма активный комплекс, стабилизируемый пропердином (фактор Р). ◊ Комплекс C3b/Bb служит С3-конвертазой и значительно повышает уровень расщепления С3 по сравнению со спонтанным. Кроме того, после связывания с C3 он расщепляет C5 до фрагментов C5a и C5b. Малые фрагменты C5a (наиболее сильный) и C3a - анафилатоксины комплемента, т.е. медиаторы воспалительной реакции. Они создают условия для миграции фагоцитов в очаг воспаления, вызывают дегрануляцию тучных клеток, сокращение гладких мышц. C5a также вызывает повышение экспрессии на фагоцитах CR1 и CR3. ◊ С C5b начинается формирование «мембраноатакующего комплекса», вызывающего перфорацию мембраны клеток микроорганизмов и их лизис. Сначала образуется комплекс C5b/C6/ C7, встраивающийся в мембрану клетки. Одна из субъединиц компонента C8 - C8b - присоединяется к комплексу и катализирует полимеризацию 10-16 молекул C9. Этот полимер и формирует неспадающуюся пору в мембране, имеющую диаметр около 10 нм. В результате клетки становятся неспособными поддерживать осмотический баланс и лизируются. | |||||||||||||||||||||||||||||||||||||||||