19. Генетические и цитологические карты хромосом.
Генетическая карта хромосомы — это схема взаимного расположения генов, находящихся в одной группе сцепления. Определение групп сцепления и расстояний между генами не является Мининым этапом построения генетической карты хромосомы, щи кильку необходимо установить также соответствие изучаемой группы сцепления определенной хромосоме. Определение группы сцепления осуществляется гибридологическим методом, т.е. путем изучения результатов скрещивания, а исследование хромосом — цитологическим методом с проведением микроско�пического исследования препаратов. Для определения соответствия данной группы сцепления конкретной хромосоме при�меняют хромосомы с измененной структурой. Выполняют стандартный анализ дигибридного скрещивания, в котором один исследуемый признак кодируется геном, локализованным на хромосоме с измененной структурой, а второй — геном, локализованным на любой другой хромосоме. В случае если наблюдается сцепленное наследование этих двух признаков, можно говорить о связи данной хромосомы с определенной группой сцепления.
Анализ генетических и цитологических карт позволил сформулировать основные положения хромосомной теории наследствен�ности.
1. Каждый ген имеет определенное постоянное место (локус)
и хромосоме.
2. Гены в хромосомах располагаются в определенной линей�ной последовательности.
3. Частота кроссинговера между генами прямо пропорциональна расстоянию между ними и обратно пропорциональна силе сцепления.
20. Кодирование и реализация биологической информации в клетке. Кодовая система ДНК и белка. Экспериментальное обоснование триплетного кода в опытах Ниринберга.
Первично все многообразие жизни обусловливается разнообразием белковых молекул, выполняющих в клетках различные биологические функции. Структура белков определяется набором и порядком расположения аминокислот в их пептидных цепях. Именно эта последовательность аминокислот в пептидных цепях зашифрована в молекулах ДНК с помощью биологического (генетического) кода. Для шифровки 20 различных аминокислот достаточное количество сочетаний нуклеотидов может обеспечить лишь триплетный код, в котором каждая аминокислота шифруется тремя стоящими рядом нуклеотидами.
Генетический код – это система записи информации о последовательности расположения аминокислот в белках с помощью последовательного расположения нуклеотидов в и-РНК.
Св-ва ген. кода:
1) Код триплетен. Это означает, что каждая из 20 аминокислот зашифрована последовательностью 3 нуклеотидов, называется триплетом или кодоном.
2) Код вырожден. Это означает, что каждая аминокислота шифруется более чем одним кодоном (исключение метиотин и триптофан)
3) Код однозначен – каждый кодон шифрует только 1 аминоксилоту
4) Между генами имеются «знаки препинания» (УАА,УАГ,УГА) каждый из которых означает прекращение синтеза и стоит в конце каждого гена.
5) Внутри гена нет знаков препинания.
6) Код универсален. Генетический код един для всех живых на земле существ.
Транскрипция – это процесс считывания информации РНК, осуществляемой и-РНК полимеразой. ДНК – носитель всей генетической информации в клетке, непосредственного участия в синтезе белков не принимает. К рибосомам – местам сборки белков – высылается из ядра несущий информационный посредник, способный пройти поры ядерной мембраны. Им является и-РНК. По принципу комплементарности она считывает с ДНК при участии фермента называемого РНК – полимеразой. В процессе транскрипции можно выделить 4 стадии:
1) Связывание РНК-полимеразы с промотором,
2) инициация – начало синтеза. Оно заключается в образовании первой фосфодиэфирной связи между АТФ и ГТФ и два нуклеотидом синтезирующей молекулы и-РНК,
3) элонгация – рост цепи РНК, т.е. последовательное присоединение нуклеотидов друг к другу в том порядке, в котором стоят комплементарные нуклеотиды в транскрибируемой ните ДНК,
4) Терминация – завершения синтеза и-РНК. Промотр – площадка для РНК-полимеразы. Оперон – часть одного гена ДНК.
ДНК (дезоксирибонуклеиновая кислота) – биологический полимер, состоящий из двух полинуклеотидных цепей, соединенных друг с другом. Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие одно из четырех азотистых оснований: аденин (А) или тимин (Т), цитозин (Ц) или гуанин (Г), пятиатомный сахар пентозу – дезоксирибозу, по имени которой получила название и сама ДНК, а также остаток фосфорной кислоты. Эти соединения носят название нуклеотидов.
21. Экспериментальные доказательства роли ДНК в передаче наследственной информации в клетке.
Опыты Херши и Чейза - белок фага метили радиоактивной серой, а ДНК радиоактивным фосфором, вновь образовавшиеся фаги содержали только радиоактивный фосфор. Опыты показали, что генетическая информация от внедрившегося фага его потомкам передается только проникающей в клетку нуклеиновой кислотой, а не белком, содержащимся в капсуле вируса. Вирусы, поражающие бактериальные клетки - бактериофаги.
Опыты Гриффитса. Он работал с двумя штаммами пневмококков. Штамм S имеет капсульную оболочку и вирулентен. При введении его мышам они погибали. Клетки штамма R не имели капсульных оболочек, гибели мышей не наступало. Клетки вирулентного штамма подвергали действию высоких температур, они не вызывали заболевания. При введении мышам смеси из невирулентного и убитого нагреванием вирулентного штаммов мыши заболели и погибли. Из крови мышей выделены живые S пневмококки. Произошла трансформация штамма R в штамм S. Трансформация - включение чужеродной ДНК в бактериальную клетку. Это перенос наследственной информации от одной клетки прокариотов к другой посредством ДНК бактерии донора или клетки донора. Поскольку клетки вирулентного штамма были убиты нагреванием, значит фактором, вызывающим трансформацию, было вещество небелковой природы. Если ДНК бактерий доноров разрушалась ферментом дезоксирибонуклазой, то трансформации не происходило. Это доказывает, что трансформация вызвана ДНК.
Хайнц Френкель-Конрад исследовал роль структурных компонентов вирусов в вирусной репликации. Чистая нуклеиновая кислота вируса табачной мозаики может заразить растение, вызывая типичную картину заболевания. Более того удалось искусственно создать вегетативные гибриды из вирусов, в которых белковый футляр принадлежал одному виду, а нуклеиновая кислота – другому. Генетическая информация гибридов всегда соответствовала тому вирусу, чья нуклеиновая кислота входила в состав гибрида. Б1Н1 (здоровый)→Б1Н2 (гибрид)→Б2Н2 (больной)
Трансдукция/перемещение - заключается в том, что вирусы, покидая бактериальные клетки, в которых паразитировали, могут захватывать с собой часть их ДНК и, перемещаясь в новые клетки, передают новым хозяевам свойства старых. Это было доказано в опытах по заражению бактерий вирусами.
22. Этапы экспрессии генов в процессе биосинтеза белка. Альтернативный сплайсинг. Регуляция этапов транскрипции и трансляции. Роль микро-РНК. Геном человека.
Экспрессия генов - программируемый геномом процесс биосинтеза белков и(или) РНК. При синтезе белков Э. г. включает транскрипцию - синтез РНК с участием ферментаРНК-полимеразы; трансляцию - синтез белка на матричной рибонуклеиновой кислоте, осуществляемый в рибосомах, и (часто) посттрансляционную модификацию белков.
Различают Э. г.: 1) конститутивную - происходящую в клетке независимо от внешних обстоятельств. Сюда относят экспрессию генов, определяющих синтез макромолекул, необходимых для жизнедеятельности всех клеток, и спец. генов (тканеспецифичная Э. г.), характерных для конкретного вида клеток. 2) Индуцибельная Э. г. определяется действием к.-л. агентов - индукторов. Ими м. б. гормоны, ростовые в-ва и в-ва, определяющие дифференцировку клеток (напр., ретиноевая к-та).
Геном человека — геном биологического вида Homo sapiens. В большинстве нормальных клеток человека содержится полный набор составляющих геном 46 хромосом: 44 из них не зависят от пола (аутосомные хромосомы), а две — X-хромосома и Y-хромосома — определяют пол (XY — у мужчин или ХХ — у женщин). В ходе выполнения проекта «Геном человека» содержимое хромосом находящихся в стадии интерфазы в клеточном ядре (вещество эухроматин), было выписано в виде последовательности символов. В настоящее время эта последовательность активно используется по всему миру в биомедицине. В ходе исследований выяснилось, что человеческий геном содержит значительно меньшее число генов, нежели ожидалось в начале проекта. Только для 1,5 % всего материала удалось выяснить функцию, остальная часть составляет так называемую мусорную ДНК[2]. В эти 1,5 % входят гены, которые кодируют РНК и белки, а также их регуляторные последовательности, интроны и, возможно, псевдогены.
23. Регуляция работы генов. Значение гистонов. Структура оперона. Роль структурных генов, промотора, оператора, регулятора, факторов транскрипции (индукторов).
Работа генов в любом организме – прокариотическом, эукариотическом, одноклеточном или многоклеточном – контролируется и координируется.
Различные гены обладают неодинаковой временной активностью. Одни из них характеризуются постоянной активностью. Такие гены отвечают за синтез белков, необходимых клетке или организму на протяжении всей жизни, например, гены, продукты которых участвуют в синтезе АТФ. Большинство же генов обладает непостоянной активностью, они работают только в определенные моменты, когда появляется необходимость в их продуктах – белках. Гены различаются и по уровням своей активности (низкий или высокий).
Белки клетки классифицируются как регуляторные и структурные. Регуляторные белки синтезируются на регуляторных генах и контролируют работу структурных генов. Структурные гены кодируют структурные белки, выполняющие структурную, ферментативную, транспортную и другие функции (кроме регуляторной!).
Регуляция синтеза белка осуществляется на всех этапах этого процесса: транскрипции, трансляции и посттрансляционной модификации либо путем индукции, либо путем репрессии.
Регуляция активности генов эукариотических организмов намного сложнее регуляции экспрессии генов прокариот, что определяется сложностью организации эукариотического организма и особенно многоклеточного. В 1961 г. французские ученые Ф. Жакоб, Ж. Моно и А. Львов сформулировали модель генетического контроля синтеза белков, катализирующих усвоение клеткой лактозы – концепцию оперона.
Опероном называют группу генов, работа которых контролируется одним геном-регулятором.
Ген-регулятор – это ген, обладающий постоянной низкой активностью, на нем синтезируется белок-репрессор – регуляторный белок, который может соединяться с оператором, инактивируя его.
Оператор – точка начала считывания генетической информации, он управляет работой структурных генов.
В структурных генах лактозного оперона содержится информация о ферментах, участвующих в метаболизме лактозы. Поэтому лактоза будет служить индуктором – агентом, инициирующим работу oпepoна.
Промотор – место прикрепления РНК-полимеразы.
Терминатор – место окончания синтеза иРНК.
При отсутствии индуктора система не функционирует, поскольку "свободный" от индуктора – лактозы – репрессор соединен с оператором. В этом случае фермент РНК-полимераза не может катализировать процесс синтеза иРНК. Если в клетке оказывается лактоза (индуктор), она, взаимодействуя с репрессором, изменяет его структуру, в результате чего репрессор освобождает оператор. РНК-полимераза соединяется с промотором, начинается синтез иРНК (транскрипция структурных генов). Затем на рибосомах формируются белки по программе иРНК-лактозного оперона. У прокариотических организмов на одну молекулу иРНК переписывается информация со всех структурных генов оперона, т.е. оперон – это единица транскрипции. Транскрипция продолжается до тех пор, пока в цитоплазме клетки сохраняются молекулы лактозы. Как только все молекулы будут клеткой переработаны, репрессор закрывает оператор, синтез иРНК прекращается.
Таким образом, синтез иРНК и, соответственно, синтез белка должны строго регулироваться, поскольку у клетки недостаточно ресурсов для одновременной транскрипции и трансляции всех структурных генов. И про-, и эукариоты постоянно синтезируют только те иРНК, которые необходимы для выполнения основных клеточных функций Экспрессия остальных структурных генов осуществляется под строгим контролем регуляторных систем, запускающих транскрипцию только в том случае, когда возникает потребность в определенном белке (белках).
24. Мультимерная организация белка на примере гемоглобина человека. Серповидно-клеточная анемия.
Гемоглоби́н (от др.-греч. αἷμα — кровь и лат. globus — шар) — сложный железосодержащий белок кровосодержащих животных, способный обратимо связываться с кислородом, обеспечивая его перенос в ткани. У позвоночных животных содержится в эритроцитах, у большинства беспозвоночных растворён в плазме крови (эритрокруорин) и может присутствовать в других тканях.
Белковая субъединица в структурной биологии — полипептид, который вместе с другими компонентами собирается в мультимерный или олигомерный белковый комплекс. Многие природные ферменты и другие белки состоят из нескольких белковых субъединиц.
Из нескольких белковых субъединиц состоят: гемоглобин, ДНК-полимеразы, нуклеосомы, мультимерными являются ионные каналы, все филаменты цитоскелета (микротрубочки, микрофиламенты и другие), рибосомы. Субъединицы таких белков могут быть идентичными, гомологичными или полностью различными, в зависимости от выполняемых функций.
В некоторых белковых комплексах одна субъединица может называться «регуляторной», а другая «каталитической». Фермент, составленный из регуляторной и каталитической субъединиц, как и фермент, составленный из главной (неактивной, апофермент) и вспомогательной (активирующей, кофермент) субъединиц часто называется холоферментом. Одна белковая субъединица представлена одной молекулой полипептида, который кодируется самостоятельным геном, таким образом, в случае сложного белка, каждой субъединице соответствует отдельный ген, либо один ген соответствует нескольким идентичным субъединицам.
Серповидноклеточная анемия — это наследственная гемоглобинопатия, связанная с таким нарушением строения белка гемоглобина, при котором он приобретает особое кристаллическое строение — так называемый гемоглобин S. Эритроциты, несущие гемоглобин S вместо нормального гемоглобина А, под микроскопом имеют характерную серпообразную форму (форму серпа), за что эта форма гемоглобинопатии и получила название серповидноклеточной анемии.
Эритроциты, несущие гемоглобин S, обладают пониженной стойкостью и пониженной кислород-транспортирующей способностью, поэтому у больных с серповидноклеточной анемией повышено разрушение эритроцитов в селезенке, укорочен срок их жизни, повышен гемолиз и часто имеются признаки хронической гипоксии (кислородной недостаточности) или хронического «перераздражения» эритроцитарного ростка костного мозга.
Серповидноклеточная анемия наследуется по аутосомно-доминантному типу (с неполным доминированием). У носителей, гетерозиготных по гену серповидноклеточной анемии, в эритроцитах присутствуют примерно в равных количествах гемоглобин S и гемоглобин А. При этом в нормальных условиях у носителей симптомы практически никогда не возникают, и серповидные эритроциты выявляются случайно при лабораторном исследовании крови. Симптомы у носителей могут появиться при гипоксии (например, при подъеме в горы) или тяжелой дегидратации организма. У гомозигот по гену серповидноклеточной анемии в крови имеются только серповидные эритроциты, несущие гемоглобин S, и болезнь протекает тяжело.
Симптомы
Усталость и анемия
Приступы боли
Отек и воспаление пальцев рук и/или ног и артрит
Бактериальные инфекции
Тромбоз крови в селезенке и печени
Легочные и сердечные травмы
Язвы на ногах
Асептический некроз
Повреждение глаз
25. Основы генетической уникальности индивидуума (иммуногенетика). Генетический комплекс гистосовместимости человека (HLA). Его значение в трансплантологии
Иммуногенетика, комплексная научная дисциплина, сочетающая методы иммунологии, молекулярной биологии и генетики для изучения наследственных факторов иммунитета, внутривидового разнообразия и наследования тканевых антигенов, генетических и популяционных аспектов взаимоотношений макро- и микроорганизма и тканевой несовместимости. Начало Иммуногенетика положили работы немецких учёных П. Эрлиха и Ю. Моргенрота, обнаруживших в начале 20 в. группы крови у коз, и открытие К. Ландштейнером групп крови у человека. Термин «Иммуногенетика» предложен американским учёным М. Ирвином в 1930.
Человеческие лейкоцитарные антигены, Система генов тканевой совместимости человека (англ. HLA, Human Leucocyte Antigens) — группа антигенов гистосовместимости, главный комплекс гистосовместимости (далее MHC) у людей. Представлены более, чем 150 антигенами. Локус, расположенный на 6-й хромосоме содержит большое количество генов, связанных с иммунной системой человека. Этими генами кодируются в том числе и антигенпредставляющие белки, расположенные на поверхности клетки. Гены HLA являются человеческой версией генов MHC многих позвоночных (на них проводилось множество исследований MHC генов).
Роли HLA важны в защите от болезней, могут быть причиной отторжения органов после пересадки, могут защищать от рака или увеличивать его вероятность (если разрегулированы из-за частых инфекций. Они могут влиять на развитие аутоиммунных заболеваний (например, сахарный диабет 1-го типа, целиакию).
В течение долгого времени в качестве идеального критерия для отбора доноров почечных аллотрансплантатов была принята совместимость по HLA-антигенам — главному генному комплексу гистосовместимости (гл. 63). Было показано, что в хромосомах млекопитающих всех изученных видов имеется единственный участок, который кодирует сильные, или главные, трансплантационные антигены. У человека имеется аналогичный 6-й хромосомный участок, называемый HLA. Тем не менее и другие антигены, называемые минорными (второстепенными), могут играть решающую роль, особенно антигены групп крови и эндотелиальный антиген, находящийся в моноцитах периферической крови, но не в лимфоцитах. Данные, указывающие на участок HLA, как на генный участок, кодирующий главные трансплантационные антигены, были получены в результате успешного приживления у реципиентов трансплантатов почек и костного мозга, взятых от доноров-родственников, причем особенно успешные результаты получались у пар донор — реципиент, представляющих собой сингенных сибсов. Тем не менее 10—15% почечных аллотрансплантатов, взятых от сингенных сибсов, часто отторгаются уже в первые недели после трансплантации. Весьма вероятно, хотя и не доказано, что подобные неудачи обусловлены предварительной сенсибилизацией к антигенам, не имеющим отношения к HLA. Антигены, не имеющие отношения к HLA, относительно слабые, и поэтому их можно подавить с помощью обычной иммуносупрессивной терапии. Однако если примирование уже произошло, то вторичные реакции будут намного более устойчивыми. На самом деле несовместимость по антигенам системы АВН представляет опасность вследствие наличия естественных анти-А и анти-В антител.
26. Структурно-функциональные уровни организации наследственного материала у прокариот и эукариот: генный, хромосомный, геномный. Ген и его свойства. Триплетный код. Внутриклеточная регуляция (гипотеза Жакоба и Моно).
Ген — структурная и функциональная единица наследственности, контролирующая развитие определённого признака или свойства. Совокупность генов родители передают потомкам во время размножения. Однако перенос генов от родителей к потомкам не является единственным способом передачи генов. В 1959 году был описан случай горизонтального переноса генов. В отличие от вертикального переноса, в горизонтальном организм передаёт гены организму, который не является его потомком. Этот способ передачи широко распространён среди одноклеточных организмов и в меньшей степени среди многоклеточных.
Свойства гена:
1. дискретность — несмешиваемость генов;
2. стабильность — способность сохранять структуру;
3. лабильность — способность многократно мутировать;
4. множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм;
5. аллельность — в генотипе диплоидных организмов только две формы гена;
6. специфичность — каждый ген кодирует свой признак;
7. плейотропия — множественный эффект гена;
8. экспрессивность — степень выраженности гена в признаке;
9. пенетрантность — частота проявления гена в фенотипе;
10. амплификация — увеличение количества копий гена.
Триплетный код — генетический код, в кото�ром каждая аминокислота полипептидной цепи определяется группой из трех нуклео�тидов ДНК.
27. Ген - функциональная единица наследственности. Молекулярное строение гена у прокариот и эукариот. Гипотеза "Один ген - один фермент", ее современная трактовка.
Все организмы на Земле подразделяют на две группы по признаку структурной организации ядра в их клетках. Те, называют
Эукариоты- многоклеточные и одноклеточные организмы, в клетках которых имеется ядро, Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой.
У эукариот ДНК линейная и связана с белками-гистонами и другими белками хромосом, которых нет у бактерий(прокариотов).
Прокариоты -одноклеточные микроорганизмы, не имеющие четко сформированного ядра, В прокариотической клетке содержится только одна хромосома, которая находится в особой области клетки — нуклеоиде, который не отделён мембраной от остальной цитоплазмы.
У прокариот ДНК кольцевая.
Организация генома прокариот (на примере кишечной палочки)
Для прокариот характерна относительно простая структура генов.
Основу генетического аппарата кишечной палочки составляет бактериальная хромосома, входящая в состав нуклеоида – ядерноподобной структуры. Нуклеоид по морфологии напоминает соцветие цветной капусты и занимает примерно 30% объема цитоплазмы. Бактериальная хромосома представляет собой кольцевую двуспиральную правозакрученную молекулу ДНК длиной около 1 мм., которая свернута во вторичную спираль. Каждая такая молекула состоит из 5-100 пар нуклеотидов. Суммарное содержание ДНК (геном) в бактериальной клетке намного меньше, чем в эукариотической, а, следовательно, меньше и объем закодированной в ней информации. В среднем такая ДНК содержит несколько тысяч генов.Длина бактериальной хромосомы составляет примерно 4,7 млн. нуклеотидных пар (п.н.), или
1,6 мм. Вторичная структура хромосомы поддерживается с помощью гистоноподобных (основных) белков и РНК. Бактериальная хромосома удваивается перед делением клетки, и сестринские копии распределяются по дочерним клеткам.Молекулы ДНК, способные себя воспроизводить путем репликации, называются репликоны.
28. Классификация генов: гены структурные, регуляторы. Свойства генов (дискретность, стабильность, лабильность, полиаллелизм, специфичность, плейотропия).
Элементарной функциональной единицей наследственного материала, определяющей возможность развития отдельного признака клетки или организма, является ген.
Структурные гены — уникальные компоненты генома, представляющие единственную последовательность, кодирующую определённый белок или некоторые виды РНК. (См. также статью гены домашнего хозяйства).
Функциональные гены — регулируют работу структурных генов.
Свойства гена
дискретность — несмешиваемость генов;
стабильность — способность сохранять структуру;
лабильность — способность многократно мутировать;
множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм;
аллельность — в генотипе диплоидных организмов только две формы гена;
специфичность — каждый ген кодирует свой признак;
плейотропия — множественный эффект гена;
экспрессивность — степень выраженности гена в признаке;
пенетрантность — частота проявления гена в фенотипе;
амплификация — увеличение количества копий гена.
29. Размножение - универсальное свойство живого, обеспечивающее материальную непрерывность в ряду поколений. Эволюция размножения, формы размножения
Размножение – это способность организмов производить себе подобных особей того же вида. Существует два типа размножения: половое и бесполое.
Древнейшим способом размножения на Земле было бесполое размножение.
Бесполое размножение.
Бесполое размножение происходит без образования специальных клеток, в нём участвует один организм, одна особь, при этом размножении образуются идентичные потомки. Единственным источником генетической изменчивости являются случайные мутации. Цитологической основой бесполого размножения является митоз. Молекулярной основой бесполого размножения является репликация ДНК. Бесполое размножение у различных живых организмов может происходить по-разному. Формы бесполого размножения:
1. Почкование – это форма бесполого размножения при которой новая особь образуется в виде выростов (почки) на теле родительской особи, а затем отделяется от неё и превращается в самостоятельную особь (гидра, дрожжи).
2. Фрагментация – это разделение особи на две или более частей, каждая из которых растёт и образуется отдельная особь (высшие растения, губка, дождевой червь).
3. Образование спор. Спора – это одноклеточная репродуктивная единица, состоящая из ядра и небольшого количества цитоплазмы под плотной оболочкой. Из споры образуется новая особь (низшие растения).
4. Деление. Бинарное деление клетки на две части. Ядро родительской особи один или несколько раз делится митозом, при этом образуется два или несколько дочерних ядер. Каждое из них окружается цитоплазмой и развивается в самостоятельный организм.
5. Шизогония – это множественное деление клетки. Сначала в клетке многократно делится ядро, затем вокруг каждого ядра обособляется участок цитоплазмы, который окружается плазматической мембраной. Затем происходит распад на отдельные клетки (малярийный плазмодий).
6. Вегетативное размножение. Осуществляется формирование дочернего организма из группы клеток материнского организма. У растений это размножение происходит за счёт вегетативных органов: корневищ, луковиц, клубней, усов.
В результате бесполого размножения образуются генетически идентичные особи. Скорость размножения очень высокая и в постоянных условиях организма быстро захватывают экологическую нишу.
Половое размножение.
Появилось половое размножение более 3 млрд. лет назад. Сущность полового размножения в перекомбинации генетического материала родительских особей. В результате дочерние особи становятся более разнообразными, и естественный отбор выбирает из них наиболее приспособленные. При половом размножении потомство получается в результате слияния гаплоидных клеток – гамет. При оплодотворении образуется зигота. Из которой развивается новый организм.
Оплодотворение – это процесс слияния сперматозоида с яйцеклеткой с последующим слиянием их ядер и образованием диплоидной зиготы. Биологическое значение этого процесса состоит в том, что при слиянии мужских и женских гамет образуется новый организм, несущий признак обоих родительских организмов.
Гаметы гаплоидны, они содержат половинный набор хромосом и образуются в результате мейоза.
Одной из модификаций полового размножения является партеногенез.
Партеногенез – это процесс, при котором женская гамета развивается в новую особь без оплодотворения (встречается у животных (пчёлы) и растений). Преимущество в том, что увеличивается скорость размножения.
30. Гаметогенез и мейоз: цитологическая и цитогенетическая характеристика. Биологическое значение мейоза. Сходства и отличия митоза и мейоза.
Гаметогенез — процесс образования яйцеклеток (овогенез) и сперматозоидов (сперматогенез) —подразделяется наряд стадий (рис. 5.4).
В стадии размножения диплоидные клетки, из которых образуются гаметы, называют сперматогониями и овогониями. Эти клетки осуществляют серию последовательных митотических делений, в результате чего их количество существенно возрастает. Сперматогонии размножаются на протяжении всего периода половой зрелости мужской особи. Размножение овогоний приурочено главным образом к периоду эмбриогенеза. У человека в женском организме этот процесс наиболее интенсивно протекает в яичниках между 2-м и 5-м месяцами внутриутробного развития. К 7-му месяцу большая часть овоцитов входит в профазу I мейоза.
Так как способом размножения клеток-предшественниц женских и мужских гамет является митоз, то овогоний и сперматогонии, как и все соматические клетки, характеризуются диплоидностью. В ходе митотического цикла их хромосомы имеют либо однонитчатую (после митоза и до завершения синтетического периода интерфазы), либо двунитчатую (постсинтетический период, профаза и метафаза митоза) структуру в зависимости от количества биспиралей ДНК. Если в одинарном, гаплоидном наборе число хромосом обозначить как п, а количество ДНК — как с, то генетическая формула клеток в стадии размножения соответствует 2п2с до S-периода и 2n4c после него.
Рис. 5.4. Схема гаметогенеза:
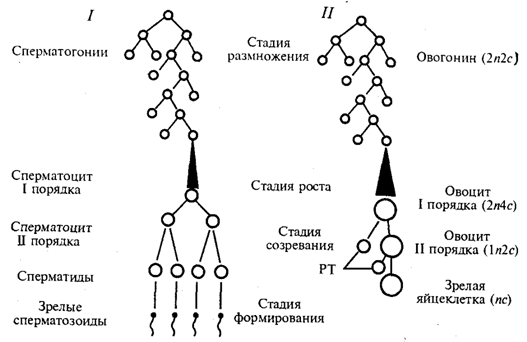
1 — сперматогенез, 2 — овогенез, n — количество хромосомных наборов,с — количество ДНК, РТ — редукционные тельца
На стадии роста происходит увеличение клеточных размеров и превращение мужских и женских половых клеток в сперматоциты и овоциты I порядка, причем последние достигают больших размеров, чем первые. Одна часть накапливаемых веществ представляет собой питательный материал (желток в овоцитах), другая — связана с последующими делениями. Важным событием этого периода является репликация ДНК при сохранении неизменным числа хромосом. Последние приобретают двунитчатую структуру, а генетическая формула сперматоцитов и овоцитов I порядка приобретает вид 2n4с.
Основными событиями стадии созревания являются два последовательных деления: редукционное и эквационное, которые вместе составляют мейоз (см. разд. 5.3.2). После первого деления образуются сперматоциты и овоциты II порядка (формула n2с), а после второго — сперматиды и зрелая яйцеклетка (пс).
В результате делений на стадии созревания каждый сперматоцит I порядка дает четыре сперматиды, тогда как каждый овоцит I порядка — одну полноценную яйцеклетку и редукционные тельца, которые в размножении не участвуют. Благодаря этому в женской гамете концентрируется максимальное количество питательного материала — желтка.
Процесс сперматогенеза завершается стадией формирования, или спермиогенеза. Ядра сперматид уплотняются вследствие сверхспирализации хромосом, которые становятся функционально инертными. Пластинчатый комплекс перемещается к одному из полюсов ядра, образуя акросомный аппарат, играющий большую роль в оплодотворении. Центриоли занимают место у противоположного полюса ядра, причем от одной из них отрастает жгутик, у основания которого в виде спирального чехлика концентрируются митохондрии. На этой стадии почти вся цитоплазма сперматиды отторгается, так что головка зрелого сперматозоида практически ее лишена.
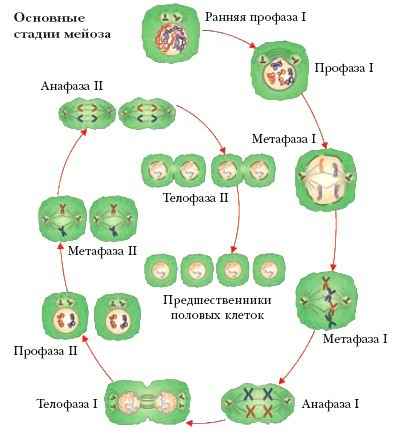
МЕЙОЗ (деления созревания, период созревания), этап в образовании половых клеток; состоит из двух последовательных делений исходной диплоидной клетки (содержат два набора хромосом – 2n) и формирования четырёх гаплоидных половых клеток, или гамет (содержат по одному набору хромосом – n). Уменьшение (редукция) числа хромосом (2nn) происходит за счёт того, что на два деления приходится лишь одно удвоение (репликация) хромосомного материала. При оплодотворении гаплоидные гаметы – яйцеклетка и сперматозоид – сливаются и диплоидное число хромосом, характерное для каждого вида, восстанавливается (n + n2n).
В главных чертах мейоз протекает сходно у разных групп организмов и у особей женского и мужского пола. Два следующих друг за другом деления первичной половой клетки обозначаются как мейоз I и мейоз II. Подобно делению соматических клеток – митозу, и мейоз I, и мейоз II состоят из четырёх основных стадий – профазы, метафазы, анафазы и телофазы. Вступающая в мейоз клетка диплоидна, а каждая хромосома содержит удвоенное количество ДНК. В первом мейотическом делении особенно сложна и длительна профаза I (у человека она занимает 22,5 сут). На этой стадии гомологичные хромосомы соединяются (конъюгируют) в пары – биваленты. В каждой хромосоме бивалента различимы в микроскопе две продольные половины – хроматиды, т. е. бивалент представляет собой четвёрку (тетраду) хроматид. В профазе I происходит генетически значимое событие – обмен гомологичными (содержащими одни и те же гены) участками несестринских хроматид, или кроссинговер. В анафазе I биваленты разъединяются и гомологичные хромосомы расходятся к противоположным полюсам клетки, причем, в отличие от анафазы митоза, каждая хромосома сохраняет две хроматиды. В результате число хромосом уменьшилось вдвое, но удвоенным остаётся и количество ДНК, представленное двумя хроматидами. Важная особенность расхождения хромосом заключается в том, что любая, отцовская или материнская, хромосома из гомологичной пары может отойти к любому из полюсов независимо от того, как расходятся хромосомы других пар. Это означает, что число возможных сочетаний хромосом в дочерних клетках обычно очень велико: 2n, где n – число хромосомных пар (у человека – 223). Так происходит ещё одно перемешивание родительского генетического материала – рекомбинация хромосом.
После мейоза I обычно сразу или после короткой интерфазы, во время которой удвоение хромосом не происходит, следует мейоз II. Это деление аналогично митозу с той разницей, что делятся гаплоидные клетки. В анафа-зе II сестринские хроматиды разделяются и, став хромосомами, расходятся к полюсам. Число хромосом и количество ДНК приходят в соответствие, и мейоз II завершается образованием четырёх гаплоидных гамет, каждая из которых несёт уникальный генетический материал. У самок, однако, лишь одна из четырёх гамет – яйцеклетка, способная к оплодотворению.
Мейоз – один из ключевых биологических процессов. Его значение состоит в поддержании в поколениях постоянства хромосомных наборов (кариотипов), т. е. в обеспечении наследственности, и в создании новых сочетаний отцовских и материнских генов, т. е. в обеспечении генотипической изменчивости.
31. Отличие овогенеза от сперматогенеза. Морфология семенников и яичников.
Гаметогенез имеет несколько стадий. Сходство сперматогенеза и овогенеза как раз и заключается в том, что три стадии у них одинаковы.
1. Стадия размножения. Первичные клетки на этой стадии называются сперматогониями и овогониями, из них в последующем образуются мужские и женские половые клетки. Половые клетки несколько раз делятся путем митоза, и количество их значительно возрастает. Сперматогонии размножаются у мужчины в течение всего репродуктивного периода, а размножение овогоний происходит в эмбриональном периоде и наиболее интенсивно происходит во 2 - 5 месяц внутриутробного развития.
2. Стадия роста. В этот период клетки значительно увеличиваются в размерах. Сперматогонии и овогонии превращаются в сперматоциты и овоциты I порядка. Овоциты I порядка достигают больших размеров, поскольку накапливают питательные вещества.
3. Стадия созревания. На этой стадии происходят два следующих друг за другом деления - мейоз I и мейоз II. После первого деления образуются сперматоциты и овоциты II порядка, а после второго деления - сперматиды и зрелые яйцеклетки с тремя полярными тельцами, которые в процессе размножения не участвуют и погибают. При созревании один сперматоцит I порядка дает четыре сперматиды, а один овоцит I порядка образует одну яйцеклетку и три полярных тельца.
Эти особенности сперматогенеза и овогенеза имеют биологический смысл, который связан с разным назначением мужских и женских гамет. Неравномерное деление клеток при овогенезе (меньше) обеспечивает формирование крупной яйцеклетки, в ней накапливается большее количество питательных веществ, так как из оплодотворенного яйца будет развиваться новый организм.
При сравнительной характеристике овогенеза и сперматогенеза можно заметить, что сперматозоидов образуется значительно больше, и это также имеет биологический смысл.
Яйцеклетку достигает только один сперматозоид, проникает в нее и доставляет свой набор хромосом. Остальные же в процессе поиска яйцеклетки массово погибают.
При сравнении овогенеза и сперматогенеза становится понятным, почему сперматозоидам нет необходимости в запасании питательных веществ - их существование кратковременно, а подвижность должна быть высокой.
4. Стадия формирования. Она характерна только для сперматогенеза. Незрелая сперматида превращается в сперматозоид, приобретая свойственный ему вид. Образование сперматозоидов у мужчин начинается только в период полового созревания и происходит в течение всего года. Период развитиясперматогоний в зрелые сперматозоиды составляет 74 дня.
Схема овогенеза и сперматогенеза
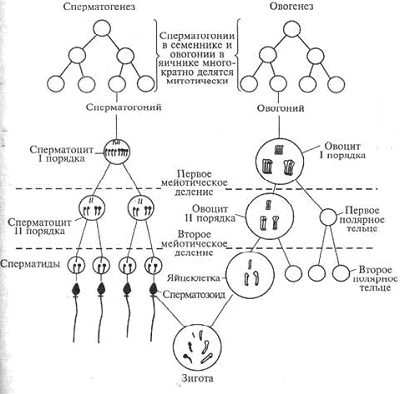
Краткую, обобщенную характеристику овогенеза и сперматогенеза представляет следующая таблица сперматогенеза и овогенеза:
признаки
|
сперматогенез
|
овогенез
|
Половые железы, половые клетки
|
Яички, сперматозоиды
|
Яичники,
яйцеклетки
|
Характерные особенности половых клеток:
- относительные размеры
- подвижность
- форма
- наличие питательных веществ
|
55 мк
подвижные
округлые со жгутиком
отсутствует
|
130-169 мк
неподвижные
округлые
имеется
|
Особенности развития половых клеток на разных стадиях:
- стадия размножения
- стадия роста
- стадия созревания
|
путем митоза образуются сперматоциты;
увеличиваются в размерах;
путем мейоза образуются гаплоидные сперматиды, их которых формируются сперматозоиды
|
путем митоза образуются овоциты;
увеличиваются в размерах;
путем мейоза образуются гаплоидные овоциты, их которых формируется яйцеклетка
|
Подытоживая все, можно привести сравнительную характеристику овогенеза и сперматогенеза. В сущности, это и будут особенности сперматогенеза и овогенеза.
1. Мы выяснили, что гаметогенез включает стадии размножения, роста и созревания клеток. Сперматогенез включает также стадию формирования (ее нет при овогенезе), в этом заключаются отличия сперматогенеза от овогенеза. Сперматозоиды проходят дополнительную четвертую стадию для того, чтобы приобрести своеобразную форму и сформировать аппарат движения.
2. Второе отличие сперматогенеза от овогенеза: из сперматоцита I порядка получается четыре половых клетки, а из ооцита I порядка получается одна полноценная половая клетка.
3. Яйцеклетки образуются циклически, процесс повторяется через 21-35 дней (менструальный цикл). После гибели яйцеклетки, что сопровождается кровотечением, изменившийся гормональный фон дает толчок к созреванию другой яйцеклетки.
Сравнительная характеристика овогенеза и сперматогенеза показывает, что у женщин мейоз начинается в период внутриутробного развития.
Ооциты I порядка у новорожденной девочки останавливаются в фазе мейоз I, и завершается созревание ооцита к моменту полового созревания. У мальчиков процесс образования сперматозоидов идет непрерывно и сохраняется в течение всей половой зрелости мужчины.
4. Из характеристики овогенеза и сперматогенеза следует, что существуют значительные различия в количестве образованных половых клеток в женском и мужском организме. Взрослый мужчина производит 30 миллионов спермиев в день, а женщина - порядка 500 зрелых яйцеклеток за всю свою жизнь.
5. Различия сперматогенеза и овогенеза заключаются также в том, что стадия размножения при сперматогенезе идет постоянно, а при овогенезе заканчивается после рождения.
6. Стадия роста при сперматогенезе короче, чем при овогенезе.
7. Стадия созревания при овогенезе имеет особенности, которые заключаются в неравномерности делений при созревании, что приводит к выделению полярных телец, что отсутствует при сперматогенезе.
8. Различия сперматогенеза и овогенеза заключаются в том, что сперматогенез более подвержен влиянию внешней среды, нежели овогенез, что связано с различием в расположении половых органов - семенники находятся вне брюшной полости.
9. Из сравнительной характеристики овогенеза и сперматогенеза можно увидеть, что, поскольку образование яйцеклеток начинается еще до рождения девочки, а завершается для яйцеклетки только после ее оплодотворения, то неблагоприятные факторы внешней среды могут повлечь генетические аномалии у потомства.
32. Характеристика основных этапов оплодотворения. Биологическое значение оплодотворения. Половой диморфизм. Партеногенез.
Оплодотворение, слияние мужской половой клетки (сперматозоида) с женской (яйцом, яйцеклеткой), приводящее к образованию зиготы — нового одноклеточного организма. Биологический смысл оплодотворения состоит в объединении ядерного материала мужской и женской гамет, что приводит к объединению отцовских и материнских генов, восстановлению диплоидного набора хромосом, а также активации яйцеклетки, то есть стимуляции её к зародышевому развитию. Соединение яйцеклетки со сперматозоидом обычно происходит в воронкообразно расширенной части маточной трубы в течение первых 12 часов после овуляции. Семенная жидкость (сперма), попадая во влагалище женщины при половом сношении (коитусе), обычно содержит от 60 до 150 млн сперматозоидов, которые благодаря движениям со скоростью 2 — 3 мм в минуту, постоянным волнообразным сокращениям матки и труб и щелочной среде уже спустя 1 — 2 минуты после полового акта достигают матки, а через 2 — 3 часа — концевых отделов маточных труб, где обычно и происходит слияние с яйцеклеткой.
Различают моноспермное (в яйцеклетку проникает один сперматозоид) и полиспермное (в яйцеклетку проникают два и более сперматозоидов, но с ядром яйцеклетки сливается только одно ядро сперматозоида) оплодотворение. Сохранению активности спермиев во время прохождения их в половых путях женщины способствует слабощелочная среда шеечного канала матки, заполненного слизистой пробкой. Во время оргазма при половом акте слизистая пробка из шеечного канала частично выталкивается, а затем вновь втягивается в него и тем самым способствует более быстрому попаданию сперматозоидов из влагалища (где в норме у здоровой женщины среда слабокислая) в более благоприятную среду шейки и полости матки. Прохождению сперматозоидов через слизистую пробку шеечного канала способствует и резко повышающаяся в дни овуляции проницаемость слизи. В остальные дни менструального цикла слизистая пробка имеет значительно меньшую проницаемость для сперматозоидов.
Многие сперматозоиды, находящиеся в половых путях женщины, могут сохранять способность к оплодотворению 48 — 72 часа (иногда даже до 4 — 5 суток). Овулировавшая яйцеклетка сохраняет жизнеспособность примерно 24 часа. Учитывая это, наиболее благоприятным временем для оплодотворения считается период разрыва созревшего фолликула с последующим рождением яйцеклетки, а также 2 — 3-й день после овуляции. Вскоре после оплодотворения начинается дробление зиготы и образование зародыша.
Партеногенез (от греч. παρθενος — девственница и γενεσις — рождение, у растений — апомиксис) — так называемое «девственное размножение», одна из форм полового размножения организмов, при которой женские половые клетки (яйцеклетки) развиваются во взрослый организм без оплодотворения. Хотя партеногенетическое размножение не предусматривает слияния мужских и женских гамет, партеногенез все равно считается половым размножением, так как организм развивается из половой клетки. Считается, что партеногенез возник в процессе эволюции организмов у раздельнополых форм.
В тех случаях, когда партеногенетические виды представлены (всегда или периодически) только самками, одно из главных биологических преимуществ партеногенеза заключается в ускорении темпа размножения вида, так как все особи подобных видов способны оставить потомство. Такой способ размножения используется некоторыми животными (хотя чаще к нему прибегают относительно примитивные организмы). В тех случаях, когда из оплодотворённых яйцеклеток развиваются самки, а из неоплодотворённых — самцы, партеногенез способствует регулированию численных соотношений полов (например, у пчёл). Часто партеногенетические виды и расы являются полиплоидными и возникают в результате отдалённой гибридизации, обнаруживая в связи с этим гетерозис и высокую жизнеспособность. Партеногенез следует относить к половому размножению и следует отличать от бесполого размножения, которое осуществляется всегда при помощи соматических органов и клеток (размножение делением, почкованием и т. п.).
|
 Скачать 0.54 Mb.
Скачать 0.54 Mb.