Ответы на экзамен по вирусологии. 1. определение вируса. Отличия и сходство с клеточными организмами. Вирусы
 Скачать 0.59 Mb. Скачать 0.59 Mb.
|

|
СТРУКТУРА ВИРУСНЫХ ЧАСТИЦ: СЕРДЦЕВИНА ВИРУСА И КАПСИД (НУКЛЕОКАПСИДЫ), ОБОЛОЧКИ ВИРИОНОВ И ИХ ПРОИСХОЖДЕНИЕ. Вирусы не имеют цитоплазмы, ядра, митохондрий и рибосом, других органелл, у многих отсутствуют ферменты. Различают простые и сложные вирусы. Простые состоят из нуклеиновой кислоты (ДНК, или РНК) и белка. Белковая оболочка, которая окружает нуклеиновую кислоту, называется капсидом. Капсид каждого вируса состоит из отдельных субъединиц – капсомеров, сложные имеют дополнительную внешнюю оболочку (суперкапсид). Нуклеокапсид оболочных вирусов окружён суперкапсидом (двойным липидным слоем с интегрированными в него белками) Источником оболочки-суперкапсида может быть цитоплазматическая мембрана, цистерны комплекса Гольджи, шероховатый эндоплазматический ретикулум клетки-хозяина Регулярно повторяющаяся морфологическая единица капсида – капсомер, его основу составляют протомеры (белковые блоки), которые состоят из полипептидных субъединиц Выступы капсомеров – пепломеры (связываются с рецепторами на поверхности клеток хозяина) Геномная нуклеиновая кислота заключена в белковый капсид, образуя нуклеокапсид / сердцевину (кор) Размер вирусов колеблется в пределах 20-400 нм ТИПЫ СИММЕТРИИ ВИРУСОВ (КУБИЧЕСКИЙ, СПИРАЛЬНЫЙ, СМЕШАННЫЙ). Кубический (он же икосаэдрический – если вирус мелкий, “сферический” – если вирус крупный). Икосаэдр сам по себе – фигура с 20ю гранями, представленными равносторонними треугольниками, 30ю рёбрами, 12ю вершинами, симметрией 2,3,5 порядков. Число треугольников в икосаэдре – триангуляционное число. Грань икосаэдра – икосадельтаэдр. Пятиугольные капсомеры – пентоны. Шестиугольные – гексоны. У разных вирусов может быть разное число капсомеров на ребре. Представители с данным типом симметрии: в. Простого герпеса, аденовирусы, в. Полиомы и др. С 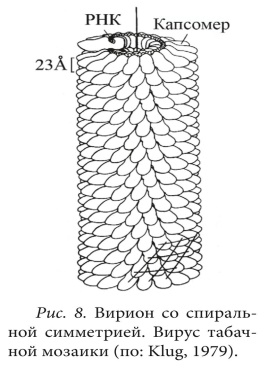 пиральный. Витки закрученной в спираль нуклеиновой кислоты окружены тесно прижатыми друг к другу капсомерами. Типичный представитель – вирус табачной мозаики (он очень устойчивый, повсеместный, заражает 125 видов растений, основной урон нанося табаку, томату, огурцу, перцу). Количественные параметры вирусов данного типа симметрии: диаметр и длина спирали, шаг спирали, число капсомеров на один оборот спирали. Смешанный. В данном случае вирион состоит из нескольких частей с разным типом симметрии, примером может послужить фаг T4 (см. рис.)  СПИРАЛЬНЫЕ ВИРУСЫ (ПРИНЦИПЫ СПИРАЛЬНОЙ СИММЕТРИИ, ВИРУС ТАБАЧНОЙ МОЗАИКИ). Витки закрученной в спираль нуклеиновой кислоты окружены тесно прижатыми друг к другу капсомерами. Количественные параметры вирусов данного типа симметрии: диаметр и длина спирали, шаг спирали, число капсомеров на один оборот спирали. Типичный представитель – вирус табачной мозаики (он очень устойчивый, повсеместный, заражает 125 видов растений, основной урон нанося табаку, томату, огурцу, перцу). Заболевание данным фитовирусом характеризуется появлением светло-зелёной пятнистости между жилками молодых листьев, далее мозаичность быстро распространяется по растению, исход хоть и не летальный, но приводит к карликовости. Иногда у яблони и винограда может проходить бессимптомно. СФЕРИЧЕСКИЕ ВИРУСЫ, ПРИНЦИПЫ ИКОСАЭДРИЧЕСКОЙ СИММЕТРИИ. Кубический тип симметрии (он же икосаэдрический – если вирус мелкий, “сферический” – если вирус крупный). Икосаэдр сам по себе – фигура с 20ю гранями, представленными равносторонними треугольниками, 30ю рёбрами, 12ю вершинами, симметрией 2,3,5 порядков. Число треугольников в икосаэдре – триангуляционное число. Грань икосаэдра – икосадельтаэдр. Пятиугольные капсомеры – пентоны. Шестиугольные – гексоны. У разных вирусов может быть разное число капсомеров на ребре. Представители с данным типом симметрии: в. Простого герпеса, аденовирусы, в. Полиомы и др. СТРОЕНИЕ НЕКОТОРЫХ СЛОЖНЫХ ВИРУСОВ (БАКТЕРИОФАГИ, ОРТО- И ПАРАМИКСОВИРУСЫ, РАБДОВИРУСЫ, РЕТРОВИРУСЫ, ВИРУС ОСПОВАКЦИНЫ, ТОГАВИРУСЫ). Б актериофаги. Ортомиксовирусы (на примере вируса гриппа). Гемагглютинин – для присоединения к клетке-хозяину, нейраминидаза – отвечает за проникновение вируса в клетку и за выход вирусных частиц после размножения. Матриксный белок М1 участвует в удерживании гемагглютинина в мембране, белок М2 участвует в образовании протонного канала). 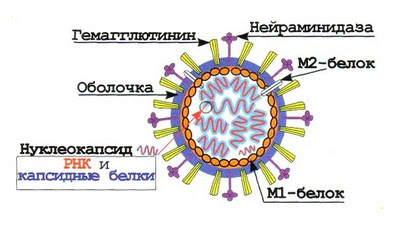 Парамиксовирусы Р 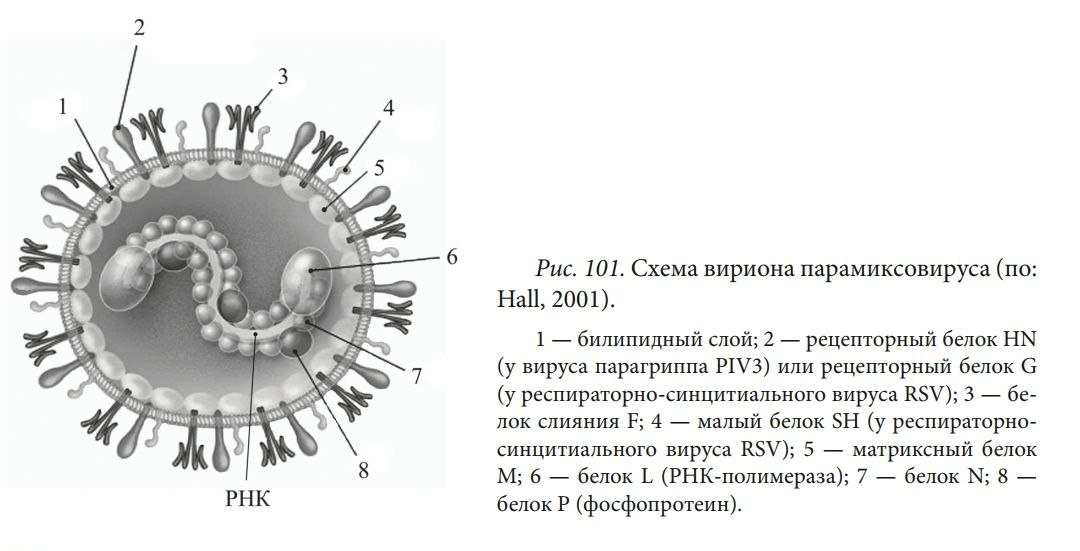 абдовирусы Р 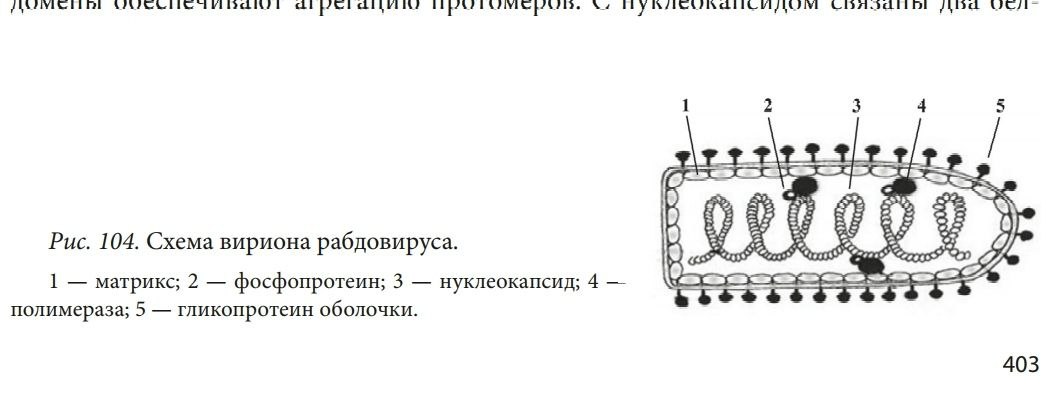 етровирусы (на примере ВИЧ), где gp – гликопротеины, а обратная транскриптаза – фермент, необходимый для синтеза ДНК и репликацию вируса в клетке. 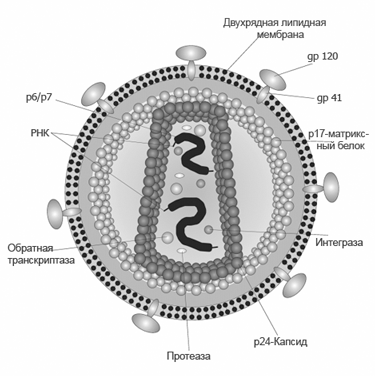 Рекомбинантный вирус осповакцины несёт трансгены противоопухолевого белка лактаптина и GM-SCF. После внутриопухолевой инъекции наблюдается ингибирование роста опухоли. Тогавирусы.  ФУНКЦИИ БЕЛКОВЫХ КОМПОНЕНТОВ ВИРИОНОВ (РЕЦЕПТОРНЫЕ ФУНКЦИИ БЕЛКОВ ВНЕШНЕЙ МЕМБРАНЫ, ФЕРМЕНТНЫЕ БЕЛКИ ВИРИОНОВ). У вирусов различают структурные и неструктурные белки. Среди структурных белков выделяют капсидные и суперкапсидные. Капсидные белки входят в состав капсомеров. Капсидные белки формируют капсид, защищая вирусную нуклеиновую кислоту. У простых вирусов капсидные белки также являются рецепторами. Суперкапсидные белки имеются у сложных вирусов. Часто это наружные гликопротеины, которые формируют шипы на поверхности суперкапсида. Они выполняют роль рецепторов – 20 узнают чувствительную клетку и адсорбируются на ней (адресные и прикрепительные белки). Для проникновения внутрь клетки (интернализации) вирусов используются матриксные М-белки и белки слияния – F-белки. F-белки обеспечивают слияние вирусной и клеточной мембран и приводят к образованию симпластов. Функции структурных белков: определяют тип симметрии вируса, обеспечивают самосборку нуклеокапсида, участвуют в распознавании клеток и взаимодействии с ними; защищают вирусный геном от нуклеаз; обладают антигенными свойствами. Среди неструктурных белков различают: – предшественники вирусных белков (обычно формируют полипротеины из которых под действием протеаз образуются конечные вирусные белки); – регуляторные белки участвуют в репродукции вируса; – вирусные ферменты: ДНК- и РНК-полимеразы обеспечивают репликацию вирусного генома; транскриптазы – синтез вирусных иРНК, интеграза вирусов выполняет встраивание вирусной ДНК в геном клетки, вирусные протеазы гидролизуют вирусные и клеточные белки и т.д. Выделяют вирионные и вирусиндуцированные ферменты вирусов. Вирионные ферменты постоянно присутствуют в составе вириона. Они участвуют в репликации, транскрипции и рекомбинации вирусных нуклеиновых кислот (эндо- и экзонуклеазы, ДНК и РНКполимеразы, обратная транскриптаза у ретровирусов и др.), адсорбции и проникновении вируса в клетку (нейраминидаза, АТФаза) и т.д. К вирусиндуцированным относятся ферменты, которые закодированы в вирусном геноме, а их синтез происходит в клетке. 17. ЛИПИДЫ И УГЛЕВОДЫ ВИРУСОВ. ДРУГИЕ КОМПОНЕНТЫ ВИРУСНЫХ ЧАСТИЦ. Липиды входят в состав суперкапсида сложных вирусов. Они включают фосфо- и гликолипиды, полученные из мембран клеток хозяина. Липиды стабилизируют вирусную частицу, определяют конформацию суперкапсидных белков, а также способствуют проникновению вируса через гидрофобную клеточную мембрану. Большинство липидсодержащих вирусов чувствительно к эфиру и детергентам. Липиды могут составлять до 20-30% от массы сложного вириона. Углеводы входят в состав гликопротеинов суперкапсида. Типичным примером такого гликопротеина является рецептор гемагглютинин, который вызывает склеивание эритроцитов и обладает антигенной специфичностью. Гликозилирование поверхностных белков влияет на их рецепторную специфичность, а также предохраняет от действия антител и клеточных протеаз. Углеводные остатки могут составлять до 10-15% от общей массы вируса. 18. ОРГАНИЗАЦИЯ ГЕНОМОВ ВИРУСОВ. ТИПЫ ДНК- И РНК-ГЕНОМОВ. ВИРУСЫ С НЕПРЕРЫВНЫМ И СЕГМЕНТИРОВАННЫМ ГЕНОМАМИ. Просто организованные вирусы представляют собой нуклеопротеиды или нуклеокапсиды и состоят из нуклеиновой кислоты (РНК или ДНК) и нескольких кодируемых ею белков, формирующих вирусную оболочку вокруг нуклеиновой кислоты — капсид. Сложно организованные вирусы содержат дополнительные оболочки, белковые или липопротеидные, и имеют более сложный химич состав. Помимо нуклеиновой кислоты и белков, они содержат липиды в наружных оболочках и углеводы в составе белков' наружных оболочек (гликопротеидов). В вирионе присутствует только один тип нуклеиновой кислоты – РНК или ДНК. И та, и другая может быть хранителем наследственной информации, выполняя функции генома. Вирусный геном может быть представлен как однонитчатыми, так и двунитчатыми молекулами РНК и ДНК. ДНК может быть линейной и кольцевой молекулой, РНК — как непрерывной, так и фрагментированной и кольцевой молекулой. Вирусные ДНК. Молекулярная масса от 1* 106 до 250* 106. В геномах, представленных двунитчатыми ДНК, информация обычно закодирована на обеих нитях ДНК: это максимальн экономия генетич материала у вирусов- свойство их как генетических паразитов. На концах молекул имеются повторы( могут быть прямыми и инвертированными). Кольцевая форма обеспечивает устойчивость ДНК к экзонуклеазам. Стадия образования кольцевой формы обязательна для процесса интеграции ДНК с клеточным геномом. В составе вирионов, содержащих однонитчатую ДНК, обычно содержатся молекулы ДНК одной полярности. Исключение составляют аденоассоциированные вирусы, вирионы которых содержат ДНК либо одной полярности (условно называемой «плюс»), либо ДНК с противоположным знаком (условно — «минус»). Поэтому тотальный препарат вируса состоит из двух типов частиц, содержащих по одной молекуле «плюс»- или «минус»-ДНК. Вирусные РНК. РНК-геном содержит около 80% вирусов. Способность РНК хранить наследственную информацию-уникальная особенность. У просто организованных и некоторых сложно организованных вирусов вирусная РНК в отсутствие белка может вызвать инфекционный процесс. Молекулы однонитчатых вирусных РНК существуют в форме одиночной полинуклеотидной цепи со спирализованными ДНК-подобными участками. При этом некомплементарные нуклеотиды, разделяющие комплементарные участки, могут выводиться из состава спирализованных участков в форме различных «петель» и «выступов». Вирусы, содержащие однонитчатые РНК, делятся на две группы. У вирусов первой группы вирусный геном обладает функциями информационной РНК, т. е. может непосредственно переносить закодированную в нем информацию на рибосомы. Вторая группа РНК-содержащих вирусов содержит геном в виде однонитчатой РНК, которая сама не обладает функциями иРНК. В этом случае функцию иРНК выполняет РНК, комплементарная геному. Синтез этой РНК (транскрипция) осуществляется в зараженной клетке на матрице геномной РНК с помощью вирусспецифического фермента — транскриптазы. У многих вирусов имеется двунитевая геномная РНК (диплорнавирусы). К ним относятся, в частности, реовирусы. Двунитевая РНК диплорнавирусов фрагментирована и состоит из нескольких сегментов. С другой стороны, и в однонитевой вирусной РНК могут образоваться двухцепочечные участки («шпильки») с поворотом цепи РНК на 180 градусов. Такие структуры принимают участие в регуляции транскрипции и трансляции вирусной РНК. 19. КОДИРУЮЩАЯ СПОСОБНОСТЬ ВИРУСНОГО ГЕНОМА Наличие сегментов ведет к увеличению кодирующей ёмкости генома. Среди РНК-геномных вирусов с одноцепочечной линейной молекулой РНК различают вирусы с +РНК (позитивным) и -РНК (негативным) геномом. +РНК выполняет и геномную и информационную функции, т. е. одновременно служит матрицей для синтеза вновь образующихся вирионных РНК и белков. Плюс-нити РНК имеют характерные окончания («шапочки») для распознавания рибосом. -РНК не способны транслировать генетическую информацию непосредственно на рибосомах, то есть они не могут функционировать как иРНК. Синтез иРНК у РНК-негативных вирусов осуществляется в зараженной клетке на матрице -РНК с помощью вирусоспецифического фермента транскриптазы. Двунитевая РНК диплорнавирусов фрагментирована и состоит из нескольких сегментов. С другой стороны, и в однонитевой вирусной РНК могут образоваться двухцепочечные участки («шпильки») с поворотом цепи РНК на 180 градусов. Такие структуры принимают участие в регуляции транскрипции и трансляции вирусной РНК. 20. ТИПЫ ВИРУСНЫХ МУТАНТОВ Вирусы-мутанты - по структуре и фенотипу отличающиеся от родительского (дикого) типа, но имеют его генетическую основу. К появлению вирусов-мутантов приводит спонтанный мутагенез, в основе которого лежит «ошибочное» спаривание азотистых оснований. Ошибки происходят потому, что в природе существует две таутомерные формы азотистых оснований. К таутомеризму ведёт постоянное перераспределение электронов и протонов в молекулах нуклеиновых кислот. Различают 4 класса вирусов-мутантов: 1) вирусы с условно дефектными геномами; 2) ДИ-частицы, т. е. дефектные интерферирующие; 3) интеграционные вирусы с дефектными геномами; 4) вирусы-сателлиты. Условно-дефектные вирусы несут мутантные геномы, дефектные в определенных условиях. Среди них чаще всего встречаются температурочувствительные ts- и холодочувствительные tc-мутанты, мутанты по спектру хозяев и мутанты по морфологии бляшек. 21. ДИ-ЧАСТИЦЫ Ди-частицы – это вирионы, у которых отсутствует некоторая часть геномной РНК или ДНК, но структурные белки остаются такими же, как у родительских вирусов. Как правило, длина нуклеокапсидов и размеры ДИ-частиц меньше родительских диких вирусов. Репликация ДИ-частиц без родительских вирионов не происходит, но при совместном заражении клеток теми и другими она восстанавливается вследствие использования генных продуктов дикого типа, которых они сами не вырабатывают. Таким образом, для ДИ-частиц родительский вирус с полноценным геномом является вирусом-помощником (хелпером). Утилизируя для своей репликации продукты генов хелпера, ДИ-частицы вместе с тем угнетают репродукцию вируса-помощника, что в вирусологии называют интерференцией, отсюда и название - дефектные-интерферирующие. 22. ГЕНЕТИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ МЕЖДУ ВИРУСАМИ Множественная реактивация. Вирусная инфекция может возникнуть при заражении клетки несколькими вирионами с поврежденными геномами вследствие того, что функцию поврежденного гена может выполнять вирус, у которого этот ген не поврежден. Вирионы с поражением разных генов дополняют друг друга путем генетической рекомбинации, в результате чего репродуцируется исходный неповрежденный вирус. Важное значение имеет расстояние между вирионами с поврежденными геномами внутри клетки. Рекомбинация. Генетической рекомбинацией называют обмен генетическим материалом, происходящий между родительскими вирусами. Возможен обмен полными генами (межгенная рекомбинация), так и участками одного и того же гена (внутригенная рекомбинация). Образующийся вирус-рекомбинант обладает свойствами, унаследованными от разных родителей.Обычно рекомбинируемые штаммы обладают характерными признаками, которые обозначаются как маркеры. Пересортировка генов. Вариантом рекомбинации является феномен, получивший название пересортировки генов. Она наблюдается при генетических взаимодействиях между вирусами, имеющими сегментированный геном. Образующиеся при этом гибридные формы вирусов называют реассортантами. Реассортанты вирусов гриппа получают при совместном культивировании вирусов с разными генами гемагглютинина и нейраминидазы. В этом случае из общего потомства путем нейтрализации соответствующих антигенов можно выделить интересующие исследователя варианты. Существуют определенные группировки (констелляции или созвездия) генов, которые в данной системе клеток более стойки и делают вирус более жизнеспособным. Перекрестная реактивация происходит в том случае, когда у одного из штаммов вируса часть генома повреждена, а другой геном интактен(неповреждённый). При смешанной инфекции двумя такими вирусами возможна рекомбинация неповрежденных участков генома инактивированного вируса с геномом интактного вируса, и в результате этого процесса появляются штаммы вируса со свойствами обоих родителей. |