Ботаника. БОТЭКЗ. 1. Особенности строения раст клетки и их отличие от жив клеток
 Скачать 1.82 Mb. Скачать 1.82 Mb.
|

|
Ботаническая номенклатура – особый раздел систематики растений. Правила использования названий регламентирует Международный кодекс ботанической литературы. В основу ботанической номенклатуры положены принципы: 1. Типификации – гербарный образец с которым связано название. 2. Приоритетности – приоритет в обнародовании (для большинства групп время отсчета – 1.05.1753 – время публикации труда Линнея «Виды растений». 3. Уникальности – каждый таксон может иметь 1 название, соответствующее Международному кодексу ботанической литературы. 4. Универсальности – названия даются на латинском языке, на других языках не считаются научными и нет правил регламентирующих эти названия. Развитие систематики как науки условно делят на следующие исторические этпы: 1. Этап прикладной систематики 2. Этап искусственной систематики 3. Этап естественной систематики 4. Этап филогенетической систематики 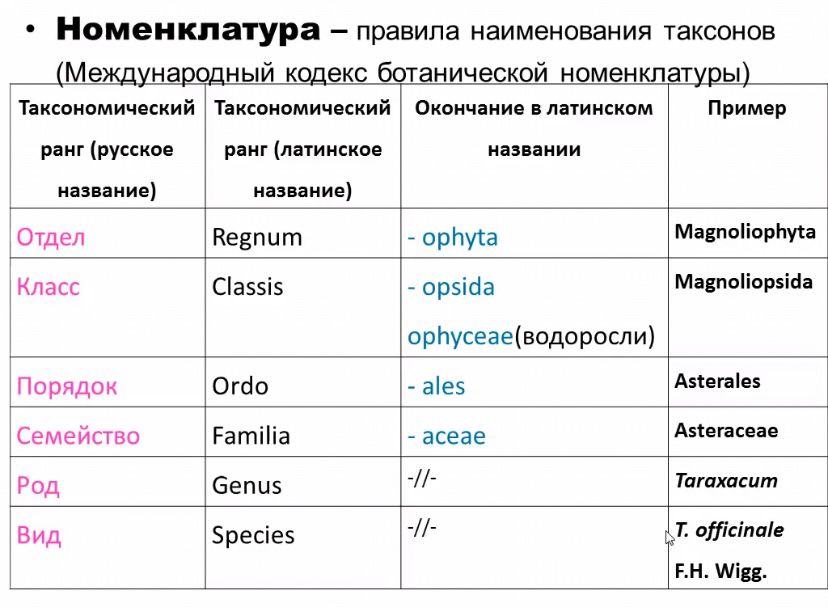  37. Отряд Риниофиты. Характеристика и систематика. Предпосылки для появления наземных растений: Ход эволюции подготовил появление более совершенных форм; За счет фотосинтеза водрослей в земной атмосфере скопилось большое кол-во кислорода; На земле в начале палеозоя происходили масштабные горнообразовательные процессы, что привело к обмелению морей и поднятию суши. Особенности отдела Тело имело вид теллома; Органы размножения- спорангии; Корневая система- ризоиды Покровная ткань- эпидерма (кутин, устьица); Равноспоровые растения Риниофиты (Rhyniophyta). Это наиболее просто устроенные высшие растения. У них был корневищеобразный орган ризоиды, от которого вверх отходили стебли( дихотомически ветвящиеся), а вниз – многочисленные р и з о и д ы.. Самые крупные представители достигали 50 см высотой и около 5 мм в диаметре. Ветви со спорангиями получили название т е л о м о в (от греч. telos – конец), а участки между разветвлениями – м е с о м а м и (от греч. mesos – средний). Листья и корни у них отсутствовали. Анатомическое строение их очень примитивное. Проводящая система была слабо развита и представляла протостелу. Наличие покровных тканей и у с т ь и ц свидетельствует о том, что это были сухопутные растения. В ископаемом состоянии гаметофит риниофитов точно не установлен. Классификация отдела риниофитов проста. Выделяют к л а с с Rhyniopsida, порядки Rhyniales и Psilophytales с соответствующими семействами. С родом псилофит сближают род тримерофит. +Риниофиты считают исходной предковой группой, от которой произошли моховидные, плауновидные, хвощевидные, псилотовидные, и папоротниковидные и праголосеменные. Они дали начало макрофилльной и микрофильной линиям эволюции высших растений. Отдел Псилотовидные (Psilotophyta) Группа близко стоящая к вымершим риниофитам. Представлены 1 классом Psilotopsida, 1 порядком Psilotales, 1 семейством Psilotaceae, 2 родами – псилот (2 вида) и тмезиптерис (10 видов). Ареал этих видов ограничен тропическими и субтропическими областями. Они являются многолетними травянистыми растениями, ведущими наземный или эпифитный образ жизни. Побеги дихотомически ветвящиеся прямостоячие, у эпифитов – свисающие. У псилота стебель плоский или трехгранный 20-100 см в высоту, у тмезиптериса высота побега 5-30 см. и они ветвятся 1 раз или вообще не ветвятся. Подземные органы как и у риниофитов называют ризомоидами. Они тоже дихотомически ветвятся и достигают длины до 1м. Ризомоиды покрыты многочисленными ризоидами. Ризомоиды могут выходить на поверхность почвы и превращаться в наземные побеги. У псилота типичные листья отсутствуют. В верхних частях стебля располагаются чешуевидные придатки без жилок. Функцию фотосинтеза выполняет стебель. У тмезиптериса в основании побега имеются чешуевидные придатки, которые выше по стеблю становятся крупнее и представляют собой низбегающие по стеблю зеленые пластинки с неветвящейся жилкой. Для надземных стеблей псилота характерно наличие эпидермы, паренхимной хлорофиллоносной коры с листовыми следами и актиностелы с 5-10 лучами ксилемы. В центре ксилемы располагается механическая ткань, выполняющая опорную функцию. У тмезиптериса, т.к. он меньших размеров в центре стебля располагается паренхима, т.е. имеет место протостела. Спорангии у псилотовых срастаются в синангии. У псилота они состоят из 3 спорангиев и имеют шаровидно-трехлопастную форму. У тмезиптериса синангий состоит из 2 спорангиев и имеет удлиненно-овальную форму. Синангии располагаются на вершинах коротких веточек, имеющих вид толстых ножек с одной чешуйкой у основания. Поэтому кажется, что синангии сидят в пазухах чешуек. Спорангии примитивные, толстостенные. Спорогенная ткань частично идет на образование спор, а частично разрушается и превращается в питательную массу. Из спор развиваются слабоветвящиеся надземные или подземные гаметофиты размером 1,5-2 см. Гаметофиты бесхлорофилльные, бурые, питаются микоризно за счет гиф гриба. Гаметофиты обоеполые, половые органы рассеяны по всей поверхности. После оплодотворения из зиготы первой развивается гаустория, остальная часть тканей зародыша делится дихотомически. Один полюс формирует надземный побег, а из другого развивается ризомоид или второй надземный побег. Систематика Отдел риниофитовые делят на 2 класса: 1. Риниевые; 2. Зоотерофилловые 38. Хар-ка класса Риниевые. Верхушечное положение спорангиев; Неравное дихотомическое ветвление; Горизонтальные и вертикальные «веточки»; Эпидерма с устьицами; Большой объем занимала кора, выполняющая ф-ию фотосинтеза; Протостела с эндархной ксилемой; Время существования: Ископаемые растения. Это древнейшая группа высших растений. Полагают, что риниофиты произошли от зеленых водорослей и первыми заселили сушу. Морфологическое строение: Большинство риниевых не имело расчленения на стебель и листья, у них не было и корней. Тело их было представлено рядом элементарных осевых участков, обладающих дихотомическим ветвлением. Эти участки получили название – телом. На конечных разветвлениях теломов развивались спорангии. Участки соединяющие теломы, называются месомами . В нижней части растения развивались теломы, прикрепляющие растение к субстрату. Они были покрыты ризоидами. Эти нижние подземные участки называют ризомоидами. Спорангии имели верхушечное (терминальное ) положение и не были еще связаны с листьями. Однако у некоторых родов уже появляются листья. Многие риниевые имели еще водорослевый облик, но другие напоминали уже плауны, хвощи, папоротники. Спорангии продолговатые или шаровидные, раскрывающиеся продольно. Стенка спорангия многослойна, покрыта кутикулой и мало отличается от наружного слоя стебля. Эпидерма стеблей покрыта довольно толстым слоем кутикулы и имеет немногочисленные устьица, что указывает на необходимость транспирации. Прикреплялись к субстрату корневищным органом ризомоидом, от которого вверх отходили стебли, а вниз многочисленные ризоиды. Риниевые были влаголюбивые растения и образовывали довольно обширные заросли на болотистых местах. Представители: Риния (Rhynia), Куксония (Cooksonia), Хорнеофит (Horneophyton). 39. Хар-ка класса зоотерофилловые Боковое положение спорангиев; Телломы стелющиеся, ветвящиеся под прямым углом; Экзархный тип протостелы; Среди зостерофилловых есть роды (Zosterophyllum, Sawdonia, Crenaticaulis, Gosslingia, Mar gophyton), объединяемые важными общими признаками и образующие вполне естественную группу. Другие роды (Nothia, Renalia) отмечены своеобразными признаками, но выделять их в особые таксоны преждевременно. Поэтому характеристика класса дана без подразделения на порядки и семейства. Для зостерофилловых характерны округлые или поперечно вытянутые спорангии с поперечной дистальной щелью, иногда сопровождаемой утолщением стенки. Спорангии раскрывались вдоль щели на две створки. Спорангии терминальные или сидят на осях сбоку на коротких ножках, собраны в верхушечные стробилы, в группы в средних частях осей, реже свободно рассеяны по побегу. Споры примерно такие же, как и у риниевых, часто с курватурами. Протоксилема экзархная, диффузно рассеяна по периферии ксилемы реже собрана в нечеткие группы. У Gosslingia, Crenaticaulis и Margophyton в месте ветвления осей есть остаток пазушной оси, возможный гомолог пазушного ризофора современной селягинеллы (в месте ветвления фертильного побега Psilophyton также есть редуцированная ось). Род Zosterophyllum имеет тонкие голые оси, ветвящиеся дихотомически, под прямым углом или К-образно. Интенсивно и неравномерно ветвящаяся часть побега, видимо, стелилась по субстрату. Спорангии большей частью собраны в терминальные стробилы (до 60 штук в одном стробиле) и прикреплялись к оси сбоку толстой ножкой, изогнутой вверх и немного вовнутрь. У Z. divaricatum спорангии спускаются по внешней стороне оси ниже ее дихотомирования. По расположению спорангиев (спиральному, двурядному, с одной стороны оси) выделяются подроды. Дистальная щель спорангия несет двугубое поперечное утолщение. Род Sawdonia установлен для покрытых эмергенцами побегов. Спорангии почковидные, с небольшими эмергенцами, дистальная щель сопровождается валиковидным утолщением. Ножка спорангия короткая, с проводящим пучком. Спорангии концентрировались небольшими группами в средних частях осей. Побеги ветвились нерегулярно, дихотомически, или выделялась главная ось. К этим двум родам близки Crenaticaulis с двумя рядами зубовидных выростов, Margophytoti с боковыми валиками на осях и Hicklingia, ранее относившаяся к риниевым. У Oricilla спорангии располагаются двумя рядами по обеим сторонам от оси и ориентированы в плоскости оси. Вероятно зостерофилловые дали начало плауновидным, причем соединительным звеном были растения типа Asteroxylon (класс Lycopodiopsida). 42. Антоцеротовые Предположительно, группа является довольно древней, возникшей еще до Девона (на это указывают разнообразные молекулярные исследования). Но наиболее древние достоверные находки ископаемых антоцеротовых (в виде спор) датируются Меловым периодом. У всех антоцеротовых гаметофит имеет вид слоевища, розетковидной или лентовидной формы. Отличаются они главным образом степенью изрезанности и волнистости краев. Слоевище стоит из нескольких слоев однотипных клеток. У большинства представителей антоцеротовых клетки имеют один пластинчатый хлоропласт с пиреноидом (это характерно для водорослей и не характерно для других растений).На нижней стороне таллома (слоевища) развиваются ризоиды и образуются межклеточные полости (вероятно, редуцированные устьица), заполненные слизью. на фотографии хорошо видны межклеточные полости В этих полостях у растений умеренных широт поселяются симбионты - сине-зеленые водоросли носток (Nostoc). Сине-зеленые водоросли осуществляют фиксацию атмосферного азота, чем существенно облегчают жизнь своим "хозяевам". Органы полового размножения развиваются эндогенно (в ткани слоевища). И только потом, при разрушении лежащих сверху клеток таллома, открываются наружу. видно, как антеридии прорываются наружу Мужские органы называются - антеридии (обычно располагаются группами), женские - архегонии. Большинство растений однодомные (антеридии и архегонии развиваются на одном растении), но могут быть и двудомными (даже представители одного вида). Спорофит образуется из оплодотворенной яйцеклетки в ткани слоевища (на месте бывшего архегония). Ткань слоевища над оплодотворенной яйцеклеткой разрастается и образует обертку вокруг молодого спорофита. Разрастаясь, спорофит прорывает обертку. Спорофит состоит из стопы, коробочки и меристематического слоя между ними. Наиболее характерным внешним признаком антоцеротовых является их длинные цилиндрические, часто слегка изогнутые растрескивающиеся коробочки роговидной формы (благодаря которым они и получили свое название: anthos (άνθος) - цветок и keros (κέρας) - рог). "Роговидные" коробочки могут достигать 16 см высотой, бывают довольно многочисленными и придают растениям своеобразный облик. При созревании коробочки сверху чернеют и растрескиваются двумя скручивающимися створками. Центральная часть коробочки бесплодна и образует колонку (развивается из эндотеция).Между колонкой и стенками коробочки развиваются тетрады спор и бесплодные нити (развиваются из амфитеция) - псевдо-элатеры. Это важный признак, отличающий антоцеротовые от других мохообразных, у которых споры развиваются из эндотеция (единственным исключением является сфагнум, у которого споры так же образуются их амфитеция). Псевдо-элатеры чувствительны к изменению влажности (гигроскопичны) и способствуют рассеиванию спор. У подавляющего большинства антоцеротовых молодая коробочка зеленая. Ее клетки имеют хлоропласты с периноидами. В наружнем слое располагаются устьица, типичного строения. Экспереминтально доказано, что спорофит антоцеротовых способен к самостоятельному питанию! Еще одной интересной особенностью антоцеротовых является практически неограниченный рост их коробочки. Коробочка растет на протяжении всей своей жизни за счет интеркалярного (вставочного) роста в меристематической зоне. Антоцеротовые неочень многочисленная группа, по разным данным в ней насчитывается от 150 до более чем 350 видов. Думаю, тут будет уместным напомнить о рангах ботанической классификации прежде, чем переходить к рассмотрению разных систем. царство (kingdom) отдел (phylum) класс (class) порядок (order) семейство (family) род (genus) вид (species) Это основные ранги, но существуют и промежуточные. Например: надкласс, подсемейство и т.п. Традиционно систематика антоцеротовых строится на макро- и микроморфологических данных. Таких, как строение спорофита, клеточная структура, ультраскульптура спор. Scanning Electron Micrographs (SEM) of hornwort spores. В верхнем ряду показаны споры двух видов рода Anthoceros, в нижнем ряду показаны вид рода Folioceros и вид рода Sphaerosporoceros. Есть четыре классификационные схемы, предложенные разными авторами в период с 1988 по 1994. Они все признают один класс антоцеротовых и расходятся в количестве признаваемых порядков 47. Общая хар-ка сосудистых споровых растений Эта группа объединяет разнообразные растения, относящиеся к 3 отделам. В жизненном цикле этих растений преобладает спорофит, который является самостоятельным растением со сложным анатомическим и морфологическим строением. Гаметофиты находятся на талломном уровне организации. Тело их состоит обычно из эпидермы и гомогенной паренхимы. Антеридии обычно сидячие, а архегонии погружены брюшком в ткань гаметофита. Сперматозоиды обычно имеют многочисленные, расположенные по спирали жгутики. Почти все современные сосудистые растения гомойгидридны (способны поддерживать относительное постоянство воды в тканях и мало зависящие от влажности окружающей среды). Низших представителей объединяют в группу сосудистых споровых растений, т.к. у них репродуктивными диаспорами являются споры. Их гаметофиты и спорофиты существуют раздельно, у многих форм гаметофиты автотрофны или микотрофны.   55. Общая характеристика семенных растений. Семенные растения – господствующая в настоящее время группа в царстве растений. Их широкое распространение связано с возникновением семени и независимостью полового процесса от капельно-жидкой влаги. У семенных растений наблюдается дальнейшая редукция гаметофита и усложнение спорофита. Гаметофит теряет свою самостоятельность, развивается не на почве, а из спор на спорофите внутри спорангиев. Все семенные растения – разноспоровые. Мегаспоры образуются в видоизмененных мегаспорангиях – семязачатках. Здесь же образуется женский гаметофит, происходит оплодотворение, и из зиготы развивается зародыш спорофита. Семязачаток превращается в семя. Семя содержит: зародыш будущего растения – спорофита со всеми основными органами – зачаточными корешком, стебельком, почечкой и зародышевыми листьями (семядолями); запас питательных веществ; защищено кожурой. Семена позволяют растениям сохранить зародыш при неблагоприятных условиях в течение довольно длительного времени. Вторая важная особенность семенных растений – возникновение в процессе эволюции пыльцевой трубки, доставляющей мужские половые клетки к архегонию и яйцеклетке. Оплодотворению предшествует опыление – перенос по воздуху пыльцевых зерен (мужских гаметофитов), в которых развиваются сперматозоиды или спермии, к семязачаткам, внутри которых формируются яйцеклетки. Семенные растения появились около 360 млн. лет назад в палеозое (конец девона). Их подразделяют на два отдела: Голосеменные – Pinophyta, или Gymnospermae и Покрытосеменные – Magnoliophyta, или Angiospermae. 56. Размножение голосеменных растений. Все голосеменные – разноспоровые растения. Имеют микро- и мегастробилы (мужские и женские шишки), различной формы, которые могут объединяться в констробилы. Микростробилы состоят из микроспорофиллов с микроспорангиями, а мегастробилы из мегаспорангиев с семязачатками. В микроспорангиях формируются микроспоры, которые начинают прорастать уже под оболочкой микроспоры. Проросшие микроспоры – мужские гаметофиты, заключенные в оболочку микроспоры называют пыльцой. После первого деления микроспоры возникает 1 маленькая клетка и 1 большая, которая может отчленить еще 1 маленькую клетку. Эти первые клетки соответствуют вегетативной части гаметофита и называются проталлиальными. Более крупная клетка – антеридиальная инициаль, делится еще раз с образованием 2 специализированных клеток. Одна из них формирует антеридий и называтся антеридиальной (спермагенной). Другая называется клеткой трубкой (вегетативной, гаусториальной). Антеридиальная клетка делится на 2 клетки. Одна остается стерильной и позже разрушается, а другая образует 2 сперматозоида и тогда называется сперматогенной, или 2 спермия и тогда называется спермагенной. Пылинки разносятся ветром и попадают на семязачаток. Развитие семязачатка начинается с появления бугорка – нуцеллуса, внутри которого выделяется 1 крупная археспориальная клетка. Она делится мейотически и 3 образуется 4 неравноценные клетки – мегаспоры. Поэтому нуцеллус рассматривают как мегаспорангий. Вслед за нуцеллусом из периферической части семязачатка формируются 2 бугорка. Которые разрастаясь и сливаясь друг с другом образуют не смыкающийся валик – интегумент. Отверстие в нем называется – микропиле, пыльцевход. Т.об., семязачаток представляет собой сложный орган, состоящий из мегаспорангия – нуцеллуса и покрова – интегумента. Три мегаспоры погибают, а 1 прорастает в женский гаметофит – эндосперм. В верхней части эндосперма образуется 2 архегония упрощенного строения. В них – по 1 яйцеклетке. Опыление происходит с помощью ветра. Пыльца попав на семяпочку прорастает. При этом трогается в рост клетка трубка (вегетативная клетка). Она растет на одном полюсе и вытягивается в пыльцевую трубку, которая растет в полость семяпочки проникая туда через микропиле и достигает архегония. Шейковые клетки архегония растворяются открывая свободный доступ к яйцеклетке. В антеридиальной клетке образуется 2 спермия. Они не имеют жгутиков и не способны к движению. После оплодотворения образуется зигота. После чего все части семяпочки претерпевают значительные изменения, в результате которых семяпочка превращается в семя. Интегумент превращается в семенную кожуру, нуцеллус расходуется на развитие зародыша и от него остается тонкая пленочка. Ткани эндосперма сильно разрастаются и в них откладываются запасные питательные вещества. Из зиготы формируется зародыш, состоящий из корешка, стебелька и почечки с 2-18 семядолями. 57. Систематика голосеменных растений. Систематика Отдел голосеменные растения (Pinophyta, Gymnospermae)- Класс семенные папоротники (Pteridospermopsida, Lyginopteridopsida) (вымершие растения)- Класс беннеттитовые (Bennettitopsida) (вымершие растения)- Класс саговниковые или цикадовые (Cycadopsida)- Класс гинкговые (Ginkgoopsida) -Класс оболочкосеменные (Chlamydospermatopsida)- Класс хвойные (Pinopsida, Coniferopsida)  58. Особенности ЖЦ, анатомического и морфологического строения, размножения класса Семенные папоротники. Жизненнвя форма- деревья, кустарники, травы; Ветвление- отсутствует или дихотомическое, боковое; укороченные побеги; Листорасположение- мутовчатое, на верхушке или по всему стеблю; Листья- голые, с волосками или железками; различные расчленения ( перистые, пильчатые, рассеченные, цельные), жилкование ( сетчатое, дуговое), форма перышек ( овальная, яйцевидная, линейная) Анатомия стебля- 3 варианта- несколько слоев вторичной ксилемы; мощная вторичная ксилема; мощная паренхима; Семязачатки одиночные или группами, окружены чашевидной купулой, срощенной с интегументом; |