Ботаника. БОТЭКЗ. 1. Особенности строения раст клетки и их отличие от жив клеток
 Скачать 1.82 Mb. Скачать 1.82 Mb.
|

|
Лизигенные возникают в результате растворения группы клеток, обособившихся внутри какой-либо ткани. Их деление приводит к образованию небольшого очага мелких клеток, вырабатывающих секрет. Впоследствии оболочки растворяются – и на их месте формируется полость, заполненная секретом. Лизигенные вместилища встречаются в кожуре цитрусовых, листьях эвкалипта. Схизогенные возникают из межклетников при отделении клеток друг от друга. При этом клетки, прилегающие к вместилищу, становятся эпителиальными, т. е. вырабатывают и выделяют в полость экскреторное вещество. Эпителий изолирует секрет от живых тканей. Схизогенные вместилища хорошо развиты у хвойных (смоляные ходы), некоторых сложноцветных, зонтичных. 3. Млечники – клетки или ряды клеток, содержащие в вакуолях млечный сок (латекс). Он может содержать смолы, каучук, эфирные масла, алкалоиды. Млечники бывают двух типов: членистые и нечленистые. Первые возникают из нескольких отдельных млечных клеток, которые в местах соприкосновения друг с другом растворяют оболочки. Их протопласты и вакуоли сливаются в единую разветвленную систему. Членистые млечники найдены у сложноцветных, маковых и многих других. Нечленистые млечники представляют собой одну гигантскую клетку, которая непрерывно растет, удлиняется и ветвится. Такими млечниками обладают молочайные, тутовые. 13. Стела и ее типы. Стела – совокупность первичных проводящих пучков вместе с заключенными между ними прочими тканями и примыкающим к коре перициклом. В 1886 году Ф. ван Тигем и А. Дулио впервые ввели понятие «стела», заложив основы стелярной теории – учения о типах строения и закономерностях эволюции центрального цилиндра всех высших растений. Наиболее древний и примитивный тип стелы – протостела – присущ первым наземным растениям (риниофитам). Последующая эволюция стелы была связана с увеличением поверхности проводящих тканей. Из протостелы развилась актиностела, ксилема которой на поперечном срезе имеет вид звезды. Данный тип стелы также характерен для примитивных растений (плауновидных, вымерших хвощевидных). Переход от протостелы к актиностеле связан с возникновением пучков, идущих в боковые органы; кроме того, в актиностеле проводящие ткани имеют большую поверхность соприкосновения с окружающими живыми тканями, что способствует лучшему проведению веществ. Результат дальнейшей специализации актиностелы – появление плектостелы, характерной для осевых органов плауновидных. В плектостеле ксилема представлена отдельными участками, расположенными параллельно или радиально друг к другу. В процессе эволюции протостела дала также начало сифоностеле. Сифоностела имеет трубчатое строение и обладает сердцевиной. Возникновение сифоностелы обеспечило существование более крупных растений, т. к. расположение ксилемы на периферии сделало стебли более прочными. Возникновение сердцевины увеличило поверхность соприкосновения проводящих тканей с живыми клетками, а также создало возможность запасания питательных веществ. Известны два типа сифоностелы: эктофлойная (флоэма охватывает ксилему снаружи) и амфифлойная (флоэма окружает ксилему снаружи и изнутри). Дальнейшее эволюционное усложнение стелы связано с развитием крупных боковых органов (вай, листьев). В местах их отхождения от стебля в стеле, в области узлов, образуются большие листовые лакуны (прорывы), заполненные паренхимой. Вследствие этого сифоностела паренхимными полосами (радиальными лучами), идущими от сердцевины к коре в радиальном направлении, рассекается на отдельные пучки, сохраняющие общее положение по цилиндрической поверхности (на поперечном срезе – кольцом). Возникают диктиостела, артростела и эустела. Диктиостела (характерна для папоротников, например для орляка, щитовника, кочедыжника) имеет вид сетчатой трубки. Она возникает из амфифлойной сифоностелы вследствие образования большого количества листовых прорывов. Эустела встречается у семенных растений и составлена из коллатеральных открытых пучков, расположенных по кругу. Артростела (характерна для хвощей) является разновидностью эустелы, представлена закрытыми пучками, расположенными вокруг центральной полости стелы и соединяющимися в узлах. Последнее звено в эволюции стелы стебля представляет собой атактостела однодольных растений. Ей присущи закрытые коллатеральные пучки, сложная картина прохождения пучков по стеблю (дугообразные индивидуальные листовые следы) с их разбросанным расположением по всему поперечному сечению стебля. 14. Основные вегетативные органы высших растений. Орган — это часть растения, имеющая определенную внешнюю (морфологическую) и внутреннюю (анатомическую) структуру в соответствии с выполняемой ею функцией. Различают вегетативные и репродуктивные органы растения. Основными вегетативными органами высших растений являются корень и побег (стебель с листьями). Они обеспечивают процессы питания, дыхания, проведения воды и растворенных в ней веществ, а также вегетативного размножения. Стебель – это осевая часть побега растений, состоящая из узлов и междоузлий и имеющая неограниченный рост. Рост стебля в длину осуществляется за счет деятельности верхушечных и вставочных меристем. Стебель несет на себе листья, почки, генеративные органы и выполняет главным образом опорную, проводящую, листонесущую функции. Иногда стебель функционирует как запасающий орган, служит для прикрепления к опоре, размножения и т. д. ЭОР Корень – осевой орган растения, обладающий, как правило, цилиндрической формой, радиальной симметрией и неограниченным ростом за счет деятельности верхушечной меристемы. Он морфологически отличается от побега тем, что обладает положительным геотропизмом (растет вниз), эндогенным ветвлением, радиальным проводящим пучком в первичном строении. Его верхушечная меристема всегда прикрыта корневым чехликом, и на нем не формируются листья и почки (за исключением придаточных). Эффективность минерального питания зависит от площади поглощающей поверхности корней, которая увеличивается благодаря обильному ветвлению, образованию корневых волосков и придаточных корней, постоянному нарастанию и передвижению всасывающих окончаний в новые участки почвы. При изучении строения молодых корешков можно выделить следующие зоны. На кончике корня располагаются клетки апикальной меристемы, обеспечивающие рост в длину и слагающие зону деления. Ее длина составляет в среднем 1–5 мм. Снаружи меристема прикрыта корневым чехликом, выполняющим защитную функцию и облегчающим продвижение корня в почве. Корневой чехлик также способствует ориентации корня в почве. За зоной деления следует зона растяжения (роста). В ней клеточные деления уже почти отсутствуют, а объем корня увеличивается за счет обводнения клеток и появления больших вакуолей. В расположенной выше зоне всасывания (поглощения) клетки прекращают рост и начинают дифференциацию. На поверхности образуются корневые волоски, выполняющие функцию всасывания воды и минеральных веществ. Эта зона непрерывно передвигается в почве по мере нарастания корневого окончания. Зона проведения обладает хорошо развитой проводящей тканью и передает почвенные растворы выше по органу. В ней также появляются боковые корни. Апикальная меристема корня откладывает клетки не только внутрь, как у побега, но и наружу, пополняя чехлик. У споровых растений (хвощи, плауны, папоротники) инициальная клетка одна. Она имеет тетраэдрическую форму и дает начало всем клеткам корня. У покрытосеменных растений инициали, как правило, расположены в три слоя. Однако строение и функционирование апикальной меристемы у однодольных и двудольных растений различно. У двудольных растений нижний ряд инициалей впоследствии дает начало корневому чехлику и ризодерме, средний слой – клеткам коры, а верхний слой образует проводящую систему. Следовательно, уже в зоне деления обнаруживается четкое разграничение на три зоны: наружную – дерматоген (производную зону нижнего ряда инициалей), среднюю – периблему (производную среднего ряда инициалей) и внутреннюю – плерому (производную верхнего слоя). У однодольных растений апекс отличается лишь тем, что инициали нижнего слоя дают начало только корневому чехлику, а ризодерма формируется из наружного слоя периблемы. Представление о происхождении тканей было заложено в теории гистогенов Дж. Ганштейна (1868). Согласно этой теории тело растения развивается из массива меристемы, состоящей из трех слоев (гистогенов): дерматогена, периблемы и плеромы. Побег – это основной орган семенных растений, выполняющий функцию воздушного питания. Состоит из оси (стебля) с расположенными на ней (нем) листьями и почками. В отличие от корня, побег расчленен на междоузлия и узлы с одним или несколькими листьями, прикрепленными к каждому узлу. Угол между стеблем и листом в месте его отхождения называют листовой пазухой. Междоузлия могут быть длинными, и тогда побег называют удлиненным; если же междоузлия короткие, побег называют укороченным. Побег, развивающийся из зародышевого стебелька, называют главным. В узлах расположены боковые почки, из которых формируются боковые побеги. Образуется система побегов. Побеги, вырастающие из почек за один вегетационный период, называют годичными. Для тропических растений характерно образование нескольких побегов за один вегетационный период. Такие побеги, образующиеся за один цикл роста несколько раз в году, называют элементарными. Нарастание побега осуществляется за счет апикальных меристем. Репродуктивные органы (спороносные колоски, стробилы или шишки, цветок, плод, семя) выполняют функции, связанные с половым и бесполым размножением растений, и обеспечивают существование вида в целом, его воспроизводство и расселение. Анатомическое строение корня также тесно связано с выполняемыми им функциями. В молодом корне можно четко проследит наличие следующих ярко выраженных зон : 1. Зона деления клеток, представляющая собой апикальную меристему, прикрытую корневым чехликом; 2. Зона роста, или растяжения, в которой клетки, уже вышедшие за пределы меристемы, находятся в разных стадиях их растяжения в продольном направлении; 3. Зона поглощения веществ и начала дифференциации постоянных тканей: ризодермы, характеризующейся развитием корневых волосков, а также проводящих элементов ксилемы и флоэмы; 4. Зона ветвления, в которой закладываются зачатки боковых корней. 15. Особенности анатомии однодольных и двудольных 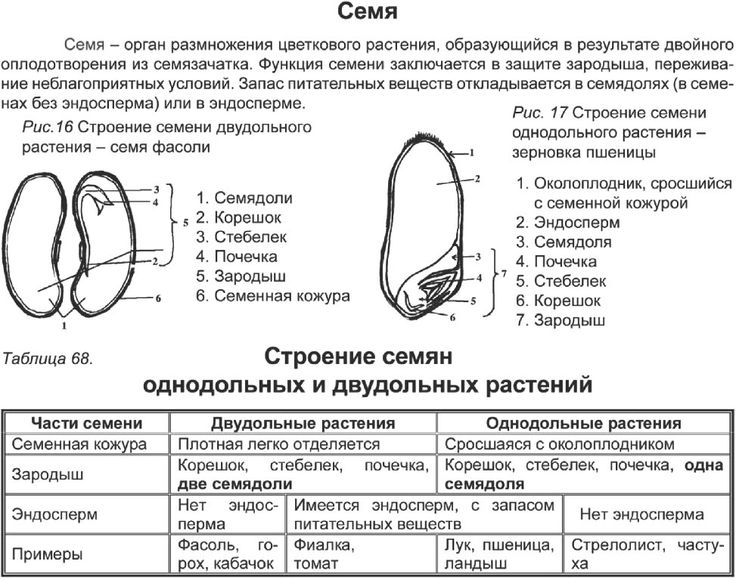 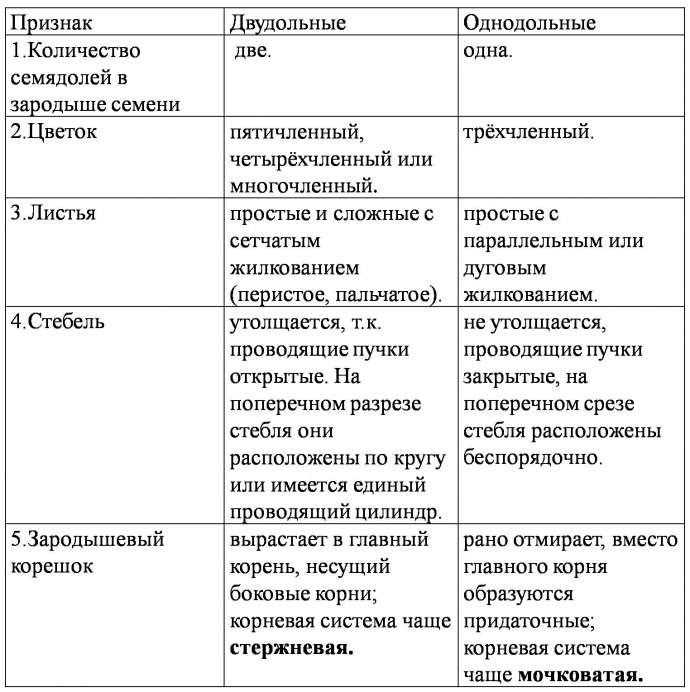 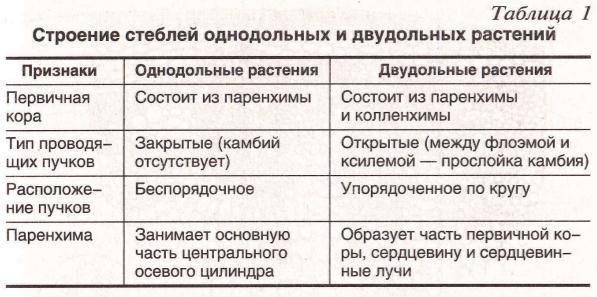 16. Первичное анатомическое строение стебля. Первичное строение стебля: в стебле, имеющем первичное строение, различают эпидермис, первичную кору и стелу (центральный цилиндр). Эпидермис располагается снаружи. В состав первичной коры могут входить ассимиляционная, механическая, запасающая, выделительная ткани, аэренхима. Стела имеет более сложное строение. Первичные проводящие ткани обычно собраны в один или несколько проводящих пучков различного типа. Расположение первичных проводящих элементов зависит от заложения и функционирования прокамбия. Если он закладывается участками, возникает пучковый тип, если сплошным слоем – непучковый. В центре стебля обычно находится сердцевина или воздушная полость (например, у тыквы, злаков). 17.Вторичное утолщение стебля для большинства двудольных и голосеменных растений характерно вторичное утолщение стеблей. Это изменение связано с деятельностью камбия и феллогена. Возникновение камбия и интенсивность его работы неодинаковы у различных растений. Различают следующие типы вторичных утолщений. 1. При пучковом типе в обособленных прокамбиальных пучках сначала возникает пучковый камбий. Затем между его разобщенными прослойками появляются перемычки межпучкового камбия. Пучковый камбий начинает откладывать проводящие элементы, а межпучковый – паренхиму или механические элементы, вследствие чего проводящие пучки остаются хорошо различимыми (Aristolochia-tip). 2. При переходном типе в прокамбиальных пучках возникает пучковый камбий, затем между проводящими пучками появляются перемычки межпучкового камбия. После этого камбий на всем протяжении начинает откладывать проводящие элементы (Ricinus-tip). У некоторых растений с ши- рокими первичными лучами отдельные участки межпучкового камбия образуют более мелкие вторичные проводящие пучки (Helianthus-tip). 3. При непучковом типе камбий закладывается как непрерывный слой (кольцо) из сплошного прокамбия и начинает откладывать на всем протяжении проводящие элементы (Saponaria-tip). У многих древесных растений первичные пучки тесно сближены, поэтому при вторичном утолщении возникают слои проводящих элементов, разделенные радиальными лучами (Tilia-tip). 18. Побег - самый изменчивый по внешнему облику орган растения. Основной тип побега зеленого растения - надземный ассимилирующий побег, несущий на оси нормальные зеленые листья. Однако наряду с фотосинтезом у побега нередко бывают и иные функции: опорная, отложение запасных веществ, вегетативное размножение, образование цветков и соцветий . Одновременно со сменой функций могут меняться облик побега и характер его роста. Пример различного характера роста дают укороченные и удлиненные побеги ( рис. 52 ). В частности, укороченные побеги имеют розеточные растения. На укороченных побегах нередко располагаются также соцветия . В ходе приспособления к иным функциям весь побег или его части могут видоизменяться - метаморфизироваться. Из частей побега чаще всего метаморфозу подвергаются листья , реже стебли , а иногда и почки . Основы теории метаморфоза растений были разработаны в конце XVIII в. великим немецким поэтом, философом и ученым И.В.Гете. Наиболее обычный случай специализации представляют собой ствол и сучья деревьев и кустарников. Известно, что их побегичерез один (у листопадных) или несколько лет (у вечнозеленых) теряют ассимиляционную функцию. Часть таких побегов иногда отмирает, но большинство остается, выполняя опорную, проводящую и запасающую функции. Из них формируется система скелетных осей, утолщающихся за счет деятельности камбия . Лишенные листьев скелетные оси известны под названием сучьев, стволов (у деревьев) и стволиков (у кустарников). Весьма обычным видоизменением побега является корневище , характерное для многих многолетних трав, полукустарников, кустарничков ( брусники , черники ) и даже кустарников (у бересклета ); луковица , резко метаморфизированный подземный, реже наземный побег с укороченным стеблем; кочан , метаморфизированная гигантская почка ( рис. 53 ). Помимо перечисленных выше видоизменений побега упомянем еще некоторые наиболее обычные его метаморфозы . Это, во-первых, колючки , которые нередко имеют стеблевое происхождение. У дикой яблони, жостера слабительного (Rhamnus cathartica) в колючки превращены укороченные побеги. Стеблевое происхождение имеют также колючки у боярышника (Crataegus) и гледичии (Gleditsia) . Образование колючек нередко связано с недостатком влаги. Виды иглицы (Ruscus) , широко распространенные в Крыму и на Кавказе, несут уплощенные пазушные побеги - филлокладии , внешне аналогичные листу ( рис. 53 ). Однако на филлокладиях образуются чешуевидные беловатые листьяи цветки , чего никогда не бывает у обычных листьев. Мелкие филлокладии сидят в пазухах чешуевидных листьев у видов спаржи (Asparagus) . 19. Корень – основной вегетативный орган высших растений, служащий для прикрепления к субстрату, поглощения из него воды и минеральных веществ. Кроме того, корень может выполнять функции запасания, синтеза различных веществ, взаимодействия с другими организмами (бактериями, грибами, корнями других растений). Корень – осевой орган растения, обладающий, как правило, цилиндрической формой, радиальной симметрией и неограниченным ростом за счет деятельности верхушечной меристемы. Он морфологически отличается от побега тем, что обладает положительным геотропизмом (растет вниз), эндогенным ветвлением, радиальным проводящим пучком в первичном строении. Его верхушечная меристема всегда прикрыта корневым чехликом, и на нем не формируются листья и почки (за исключением придаточных). Эффективность минерального питания зависит от площади поглощающей поверхности корней, которая увеличивается благодаря обильному ветвлению, образованию корневых волосков и придаточных корней, постоянному нарастанию и передвижению всасывающих окончаний в новые участки почвы. При изучении строения молодых корешков можно выделить следующие зоны. На кончике корня располагаются клетки апикальной меристемы, обеспечивающие рост в длину и слагающие зону деления. Ее длина составляет в среднем 1–5 мм. Снаружи меристема прикрыта корневым чехликом, выполняющим защитную функцию и облегчающим продвижение корня в почве. Корневой чехлик также способствует ориентации корня в почве. За зоной деления следует зона растяжения (роста). В ней клеточные деления уже почти отсутствуют, а объем корня увеличивается за счет обводнения клеток и появления больших вакуолей. В расположенной выше зоне всасывания (поглощения) клетки прекращают рост и начинают дифференциацию. На поверхности образуются корневые волоски, выполняющие функцию всасывания воды и минеральных веществ. Эта зона непрерывно передвигается в почве по мере нарастания корневого окончания. Зона проведения обладает хорошо развитой проводящей тканью и передает почвенные растворы выше по органу. В ней также появляются боковые корни. Апикальная меристема корня откладывает клетки не только внутрь, как у побега, но и наружу, пополняя чехлик. У споровых растений (хвощи, плауны, папоротники) инициальная клетка одна. Она имеет тетраэдрическую форму и дает начало всем клеткам корня. У покрытосеменных растений инициали, как правило, расположены в три слоя. Однако строение и функционирование апикальной меристемы у однодольных и двудольных растений различно. У двудольных растений нижний ряд инициалей впоследствии дает начало корневому чехлику и ризодерме, средний слой – клеткам коры, а верхний слой образует проводящую систему. Следовательно, уже в зоне деления обнаруживается четкое разграничение на три зоны: наружную – дерматоген (производную зону нижнего ряда инициалей), среднюю – периблему (производную среднего ряда инициалей) и внутреннюю – плерому (производную верхнего слоя). У однодольных растений апекс отличается лишь тем, что инициали нижнего слоя дают начало только корневому чехлику, а ризодерма формируется из наружного слоя периблемы. Представление о происхождении тканей было заложено в теории гистогенов Дж. Ганштейна (1868). Согласно этой теории тело растения развивается из массива меристемы, состоящей из трех слоев (гистогенов): дерматогена, периблемы и плеромы. Корень — осевой орган, обладающий способностью к неограниченному росту и свойством положительного геотропизма. Функции корня: • укрепление растения в почве и удержание надземной части растения; • поглощение воды и минеральных веществ; • проведение веществ; • может служить местом накопления питательных веществ; • служит органом вегетативного размножения. 20.Первичное строение корня Рассмотрим поперечный срез корешка, сделанный в зоне всасывания. Такое первичное строение корня характерно большинству семенных растений. Снаружи корня расположена ризодерма – всасывающая ткань, через которую происходит поглощение воды и минеральных веществ, а также взаимодействие с живым населением почвы. В ней различают два типа клеток: трихобласты, образующие корневые волоски, и атрихобласты. Под ризодермой находится первичная кора, выполняющая защитную, проводящую, запасающую и другие функции. Наружный слой первичной коры после отмирания корневых волосков и сбрасывания ризодермы дифференцируется в экзодерму, а из внутреннего слоя развивается эндодерма. Эндодерма в своем развитии может пройти три ступени. На первой ее клетки расположены плотно, без межклетников, и характеризуются наличием на радиальных и поперечных стенках утолщений в виде рамочек, опоясывающих клетки (поясков Каспари), в которых откладываются гидрофобное вещество, сходное с суберином, и лигнин. Пояски соседних клеток смыкаются – и создается их непрерывная цепь вокруг стелы. Пояски Каспари непроницаемы для растворов; следовательно, вещества из коры в стелу и обратно могут пройти только по симпласту, т.е. через живые протопласты клеток эндодермы. Таким образом, функция эндодермы – контроль за проведением веществ. У многих растений эндодерма получает вторичное и третичное развитие. На второй ступени развития субериноподобные вещества откладываются по всей внутренней поверхности стенок эндодермы. Однако при этом эндодерма не становится абсолютно непроницаемой для растворов, поскольку в ней имеются пропускные клетки, сохранившие первичное строение. Третья ступень характеризуется сильным утолщением и одревеснением боковых и внутренних стенок и отмиранием протопластов. Пропускные клетки сохраняются и осуществляют физиологическую связь между первичной корой и центральным цилиндром. В центре корня располагается центральный цилиндр (стела). Он имеет следующее строение. Сразу под эндодермой находится перицикл – ткань, образующая боковые корни. Клетки прокамбия формируют первичные проводящие элементы. Флоэма начинает развиваться раньше ксилемы. Клетки протофлоэмы примыкают к перициклу. Метафлоэма расположена ближе к центру. Ксилема формируется позже. Протоксилема возникает вплотную к перициклу, метаксилема образуется ближе к центру корня, т. е. ксилема и флоэма в корне закладываются экзархно и развиваются центростремительно. На поперечном срезе ксилема выглядит, как звезда, число лучей которой – от одной (папоротники) до тридцати. Радиальные пучки, имеющие по одному тяжу ксилемы и флоэмы, называют моноархными, по два – диархными, по три – триархными, по нескольку – полиархными. Между этими лучами располагаются участки флоэмы. Такой тип проводящей системы облегчает поступление растворов в сосуды, т.к. лучи ксилемы подходят к периферии стелы. 21.Вторичное утолщение корня У двудольных и голосеменных растений наблюдается вторичное утолщение корня, связанное с деятельностью камбия и феллогена. Оно начинается с закладки участков камбия между лучами флоэмы и ксилемы. Эти участки имеют вид вогнутых внутрь дуг, растущих в направлении перицикла. Когда камбиальные дуги достигают перицикла, его клетки тоже начинают делиться, соединяя отдельные дуги и образуя сплошной слой латеральной меристемы. Клетки перицикла становятся камбиальными и впоследствии дают начало радиальным лучам. Камбий откладывает внутрь вторичную ксилему, наружу – вторичную флоэму. Феллоген образуется из производных клеток перицикла и начинает откладывать перидерму. После формирования пробки первичная кора, лишенная связи с центральным цилин дром, отмирает и под давлением нарастающей изнутри массы сбрасывается. Перидерма становится покровной тканью корня. Для корней, выполняющих запасающую функцию, возможны отклонения от нормального вторичного утолщения. Например, у свеклы разрастание в толщину осуществляется за счет аномального роста (закладывается не один, а несколько слоев камбия). Добавочные слои возникают снаружи центрального цилиндра, вследствие чего образуется несколько колец проводящих элементов и запасающей паренхимы. Между ними расположены тяжи радиальной паренхимы. |