нор.физ. нор. 1. Павлов, Сеченов, Анохин, Овсянников и тд, вклад в физиологию
 Скачать 1.37 Mb. Скачать 1.37 Mb.
|

|
Функциональная система поведенчиского акта – совокупность различных нервных центров, временно объеденных для достижения цели. Как только эта цель достигнута, функц. Система распадается ,а когда появляется новая цель – формируется новая функциональная система. 1.Афферентный синтез – анализ и сопостановление различной информации , необходимой для принятия решения. На этой стадии объединяются : - пусковая афферентация – внешний раздражитель, который запускает переход к выполнению следующей стадии работы функциональной системы. - обстановочная афферентация – информацию о тех внешних условиях , в которых в данный момент находится организм. - доминирующая мотивация – информация о текущих потребностях. - память – общий опыт. 2. Принятие решения о целесообразном действии, после чего одновременно формируются : - программа будущего действия - акцептор результата действия – нейроная модель ( прообраза) параметров ожидаемого результата действия 3. Совершение дейстия и получения его результата. 4. Обратная афферентация – получение информации о параметре результата действия 5. Сравнительные параметры полученного результата с ожидаемым. В случаи совпадения этих параметров функциональная система распадается , т.к. выполнила свою задачу . Это сопровождается + эмоцией . При не совпаданиее будет – эмоция, затем вносится коррекция в программу и действие повторяется до получения планированного результата. 2.Биоритмы и их физиологическое значение. Классификация, значение в клинической практике. Биоритмы Биоритмами называются циклические изменения функций органов, систем и организма в целом. Главной характеристикой циклической активности является ее периодичность, т.е. время за которое совершается один полный цикл. В соответствии с временем периодов циклической активности все биологические ритмы делятся на три группы: 1.Циркадианные. Это ритмы с суточной периодичностью. К циркадианным ритмам относятся циклы сон-бодрствование, циклические суточные колебания температуры тела, содержания электролитов в жидкостях организма, уровня гормонов в крови и т.д.. 2.Инфрадианные. Циклы с большей, чем сутки, длительностью. Это недельные, месячные, годовые ритмы. Ими являются менструальный цикл, сезонные изменения функций организма, жизненный цикл. 3.Ультрадианные. Ритмы с периодичность меньше суток. Выделяют часовые, минутные, секундные ритмы. К этой группе биоритмов относятся потребление пищи, периодическая деятельность органов пищеварения, дыхание, сердечный ритм, ритмы активности различных клеток-пейсмекеров и т.д.. При быстром изменении параметров ритмичности геофизических процессов (например продолжительности светового дня, времени суток) нормальная циркадианная ритмичность нарушается. Возникает десинхроноз. Например состояние десинхроноза возникает у людей в связи с профессиональной деятельность (люди работающие посменно). Десинхроноз приводит к обострению хронических и возникновению других заболеваний. 3Физиология желчи.Желчеобразованиеи желчвыведение Состав желчи. Желчь является не только секретом, но и экскретом. Вместе с желчью из организма выводятся различные эндогенные и экзоген ные вещества. В желчи содержатся также белки, аминокислоты, витамины и другие вещества. Желчь обладает небольшой ферментативной активнос тью; рН печеночной желчи 7,3—8,0. Качественное своеобразие желчи определяют находящиеся в ней желч ные кислоты, пигменты и холестерин. Основное количество желчных кислот и их солей содержится в желчи в виде соединений с гликоколом и таурином .При употребле нии пищи, богатой углеводами, в желчи увеличивается содержание глико-холевых кислот, при высокобелковой диете — таурохолевых. Желчные кис лоты и их соли определяют основные свойства желчи как пищеварительно го секрета. Функции желчи. 1. Участвует в процессах пищеварения: • эмульгирует жиры, тем сам увеличивает поверхность для гидролиза их липазой; • растворяет продукты гидролиза жира, чем способствует их всасыванию; • повышает активность ферментов (панкреатических и кишечных), особенно липаз; • нейтрализует кислое желудочное содержимое; • инактивирует пепсины; • способствует всасыванию жирорастворимых витаминов, холестерина, аминокислот и солей кальция; • участвует в пристеночном пищеварении, облегчая фиксацию ферментов; • усиливает моторную и секреторную функцию тонкой кишки. 2. Стимулирует желчеобразование и желчевыделение. 3. Участвует в печеночно-кишечном кругообороте компонентов желчи - компоненты желчи поступают в кишечник, всасываются в кровь, включаются вновь в состав желчи. 4. Желчь обладает бактериостатическим действием - тормозит развитие микробов, предупреждает развитие гнилостных процессов в кишечнике. Динамика желчеобразования и желчевыделения. У человека за сутки об разуется около 500—1500 мл желчи. Процесс образования желчи (желчеотделение, или холерез) идет непрерыв но, а поступление желчи в двенадцатиперстную кишку (желчевыделение, или холекинез) — периодически, в основном в связи с приемом пищи. Нато щак желчь в кишечник поступает в соответствии с голодной периодической деятельностью. В периоды покоя она направляется в желчный пузырь, где при депонировании концентрируется и несколько изменяет свой состав. Поэтому принято говорить о двух видах желчи — печеночной и пузырной. Регуляция желчеобразования. Образование желчи происходит путем активного и пассивного транс порта веществ из крови через клетки и межклеточные контакты (вода, глю коза, креатинин, электролиты, витамины, гормоны и др.), активной секре ции компонентов желчи (желчные кислоты) гепатоцитами и обратного вса сывания воды и ряда веществ из желчных капилляров, протоков и желчно го пузыря. Ведущая роль в образовании желчи принадлежит секреции. Желчеобразование идет непрерывно, но интенсивность его изменяется за счет регуляторных влияний. Усиливают желчеобразование акт еды, при нятая пища. Рефлекторно желчеобразование изменяется при раздражении интерорецепторов желудочно-кишечного тракта, внутренних органов и ус-ловнорефлекторном воздействии. Вегетативная регуляция. Парасимпатические холинергические нервные во локна усиливают, а симпатические адренергические — снижают желчеобразо вание. Имеются экспериментальные данные и об усилении желчеобразова ния под влиянием симпатических нервов. Гуморальная регуляция. К числу гуморальных стимуляторов желчеобра зования (холеретиков) относится сама желчь. Чем больше желчных кислот поступает из тонкой кишки в портальный кровоток, тем больше их выделя ется в составе желчи, меньше желчных кислот синтезируется гепатоцитами. Если в портальный кровоток поступает мало желчных кислот, их дефицит восполняется усилением синтеза желчных кислот в печени. Секретин уси ливает секрецию желчи, выделение в ее составе воды и электролитов. Сла бее стимулируют желчеобразование глюкагон, гастрин и холецистокинин. Регуляция желчевыведения. Движение желчи в желчевыделительном аппарате обусловлено разнос тью давления в его частях и в двенадцатиперстной кишке, состоянием сфинктеров и внепеченочных желчных путей, сокращениями гладких мышц протоков и желчного пузыря. Эти сокращения согласованы с тону сом сфинктеров и регулируются нервными и гуморальными механизмами. Давление в общем желчном протоке колеблется от 4 до 300 мм вод.ст. Вне пищеварения в желчном пузыре давление составляет 60—185 мм вод.ст., во время пищеварения за счет сокращения пузыря оно возрастает до 150— 260 мм вод.ст., обеспечивая выход желчи в двенадцатиперстную кишку через открывающийся сфинктер Одди. Вид, запах пищи, подготовка к ее приему и сам приемвызывают сложное и неоднозначное у разных лиц изменение желчевыделения. Желчный пу зырь при этом через различный латентный период сначала расслабляется, а затем сокращается. Через сфинктер Одди небольшое количество желчи вы ходит в двенадцатиперстную кишку. Этот период первичной реак ции желчевыделительного аппарата длится 7—10 мин. На смену ему при ходит основной — эвакуаторный период (или период опорож нения желчного пузыря), во время которого сокращения желчного пузыря чередуются с расслаблением и через открытый сфинктер Одди в двенадца типерстную кишку переходит сначала в основном пузырная желчь, а в по следующем — печеночная. Сильными возбудителями желчевыделения являются яичные желтки, молоко, мясо и жиры. Рефлекторная стимуляцияжелчевыделительного аппарата и холекинеза осуществляется условно- и безусловнорефлекторно с рецепторов ротовой полости, желудка и двенадцатиперстной кишки через посредство блуждаю щих нервов. Гуморальная регуляция, а именно усиление желчевыделения, наблюдает ся у голодных собак при введении им крови накормленных животных. Ос новным гуморальным стимулятором желчевыделения является холецисто-кинин, вызывающий сокращение желчного пузыря. Слабые сокращения желчного пузыря возникают под влиянием гастрина, секретина, бомбезина. Тормозят сокращение желчного пузыря глюкагон, кальцитонин, антихоле-цистокинин, вазоинтестинальный пептид, панкреатический полипептид.
При необходимости быстро увеличивается мок при использовании конечного систолического (резервного объема) Билет 51. 1. женские половые гормоны. Эстрогены синтезируются фолликулами яичников. В класс эстрогенов входят: эстрон, эстрадиол, эстриол, эстерон, эстроген. Благодаря эстрогенам в пубертантном периоде происходит формирование органов по женскому типу, а так же они стимулируют развитие молочных желез. Усиливают синтез белка в печени, участвуют в синтезе альбуминов. Активируют синтез вазопрессина и окситоцина, влияют на развитие костного скелета, тормозят рост костей в длину, усиливают образование жира и его распределение, типичное для женской фигуры, способствуют оволосению по женскому типу, влияют на эмоциональное состояние, способствуют росту мышечной ткани матки во время беременности. Прогестерон синтезируется желтым телом яичников. Подготавливает эндометрий к имплантации оплодотворенной яйцеклетки. Усиливает пролиферацию и секреторную активность клеток эндометрия и молочных желез, в цитоплазме накапливаются липиды и гликоген, усиливается васкуляризация. Прогестерон тормозит продукцию ФСГ Продукция эстрогенов и прогестерона регулируется гонадотропными гормонами аденогипофиза, выработка которых увеличивается у девочек в возрасте 9-10 лет. При высоком содержании в крови эстрогенов угнетается секреция ФСГ и лютеинизирующего гормона аденогипофизом, а также гонадолиберина гипоталамусом. Менструальный цикл — это изменения слизистой оболочки матки, целью которых является подготовка наиболее благоприят ных условий для имплантации зародыша, а при ее отсутствии происходит отторжение эпителия. Фазы: Менструальный цикл включает четыре периода. 1. Предовуляционный (с 5 по 14 день). Изменения обусловлены действием ФСГ, в яичниках происходит усиленное образование эстрогенов, они стимулируют рост матки, разрастание слизистой оболочки и ее желез, ускоряется созревание фолликула, поверхность его разрывается, и из него выходит яйцеклетка – происходит овуляция. 2. Овуляционный (с 15 по 28 день). Начинается с выхода яйцеклетки в трубу, сокращение гладкой мускулатуры трубы способствует продвижению ее к матке, здесь может произойти оплодотворение. Оплодотворенное яйцо, попадая в матку, прикрепляется к ее слизистой и наступает беременность. Если оплодотворение не произошло, наступает послеовуляционный период. На месте фолликула развивается желтое тело, оно вырабатывает прогестерон. 3. Послеовуляционный период. Неоплодотворенное яйцо, достигая матки, погибает. Прогестерон уменьшает образование ФСГ и снижает продукцию эстрогенов. Изменения, возникшие в половых органах женщины исчезают. Параллельно уменьшается образование лютропина, что ведет к атрофии желтого тела. За счет уменьшения эстрогенов матка сокращается, происходит отторжение слизистой оболочки. В дальнейшем происходит ее регенерация. 4. Период покоя и послеовуляционный период продолжаются с 1 по 5 день полового цикла 2. сравнение пд в сердечной, п/п мускулатуре и нервной ткани В нервной ткани потенциал действия, как правило, возникает при деполяризации — если деполяризация мембраны нейрона достигает некоторого порогового уровня или превышает его, клетка возбуждается, и от её тела к аксонам и дендритам распространяется волна электрического сигнала. (В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы, которые сильно отличаются от потенциала действия по своей природе — например, они не подчиняются принципу «всё или ничего». Эти потенциалы преобразуются в потенциал действия на особом участке мембраны — аксонном холмике, так что потенциал действия не распространяется на дендриты). Это обусловлено тем, что на мембране клетки находятся ионные каналы — белковые молекулы, образующие в мембране поры, через которые ионы могут проходить с внутренней стороны мембраны на наружную и наоборот. Большинство каналов ионоспецифичны — натриевый канал пропускает практически только ионы натрия и не пропускает другие (это явление называют селективностью). Мембрана клеток возбудимых тканей (нервной и мышечной) содержит большое количество потенциал-зависимых ионных каналов, способных быстро реагировать на смещение мембранного потенциала. Деполяризация мембраны в первую очередь вызывает открытие потенциал-зависимых натриевых каналов. Когда одновременно открывается достаточно много натриевых каналов, положительно заряженные ионы натрия устремляются через них на внутреннюю сторону мембраны. Движущая сила в данном случае обеспечивается градиентом концентрации (с внешней стороны мембраны находится намного больше положительно заряженных ионов натрия, чем внутри клетки) и отрицательным зарядом внутренней стороны мембраны (см. Рис. 2). Поток ионов натрия вызывает ещё бо́льшее и очень быстрое изменение мембранного потенциала, которое и называют потенциалом действия (в специальной литературе обозначается ПД). 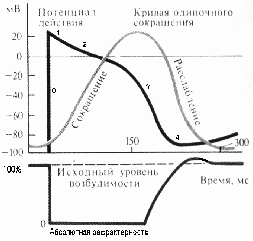 Графики сокращения, потенциала действия и возбудимости миокарда желудочков. Потенциал действия клеток рабочего миокарда состоит из фазы быстрой деполяризации, начальной быстрой реполяризации, переходящей в фазу медленной реполяризации (фаза плато) и фазы быстрой конечной реполяризации (рис. 1). Фаза быстрой деполяризации создается резким повышением проницаемости мембраны для ионов натрия, что приводит к возникновению быстрого входящего натриевого тока. Последний, однако, при достижении мембранного потенциала 30–40 мВ, инактивируется и в последующем, вплоть до инверсии потенциала (около +30 мВ) и в фазу «плато», ведущее значение имеют кальциевые ионные токи. Деполяризация мембраны вызывает активацию кальциевых каналов, в результате чего возникает дополнительный деполяризующий входящий кальциевый ток. Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для кальция и повышением проницаемости для калия. В результате входящий ток кальция уменьшается, а выходящий ток калия возрастает, что обеспечивает быстрое восстановление мембранного потенциала покоя. Длительность потенциала действия кардиомиоцитов составляет 300–400 мс, что соответствует длительности сокращения миокарда . Электрохимический этап мышечного сокращения. 1. Генерация потенциала действия. Передача возбуждения на мышечное волокно происходит с помощью ацетилхолина. Взаимодействие ацетилхолина (АХ) с холинорецепторами приводит к их активации и появлению потенциала действия, что является первым этапом мышечного сокращения. 2. Распространение потенциала действия. Потенциал действия распространяется внутрь мышечного волокна по поперечной системе трубочек, которая является связывающим звеном между поверхностной мембраной и сократительным аппаратом мышечного волокна. 3. Электрическая стимуляция места контакта приводит к активации фермента и образованию инозилтрифосфата, который активирует кальциевые каналы мембран, что приводит к выходу ионов Ca и повышению их внутриклеточной концентрации. 3.белки их переваривание Всасывание белков наиболее интенсивно протекает в верхних отделах тонкого кишечника, причем белки животного происхождения составляют 90–95 %, а растительного – 60–70 %. Основными продуктами распада, которые образуются в результате обмена веществ, являются аминокислоты, полипептиды, пептоны. Для транспорта аминокислот необходимо наличие молекул переносчика. Выделено четыре группы транспортных белков, обеспечивающих активный процесс всасывания. Поглощение полипептидов происходит пассивно по градиенту концентрации. Продукты поступают непосредственно во внутреннюю среду и с током крови разносятся по организму. Белки в пищеварительный тракт поступают с пищей, с пищеварительными соками и извлекаются из отторгнутых энтероцитов. В результате их гидролиза, с помощью различных пептидаз образуются ди-, трипептиды, аминокислоты. Основная часть белков всасывается в тонком кишечнике и только 10% всасывается в толстой кишке. Ди- и трипептиды поступают в клетку активным или пассивным путем и в цитозоле подвергаются гидролизу ферментами клетки. Аминокислоты, образуемые на мембране щеточной каймы, сразу поступают на транспортный конвейер. Существуют 4 основные транспортные системы: для нейтральных, двуосновных, дикарбоновых аминокислот и для иминокислот. Транспорт аминокислот нартийзависимый. Аминокислоты одной группы конкурируют за переносчик, поэтому если концентрация какой-либо аминокислоты находится за пределами насыщения - другие аминокислоты могут не всосаться. Белки пищи существенно различаются по аминокислотному составу. В связи с этим введено понятие биологической ценности белков пищи. Белки, содержащие весь необходимый набор аминокислот (по составу и количеству), который обеспечивает нормальные процессы синтеза, являются биологически полноценными белками. Белки, не содержащие каких-либо аминокислот, особенно незаменимых, являются неполноценными. О характере белкового обмена судят по азотистому балансу. уклеозиды транспортируются так же как и аминокислоты, специфическими переносчиками. |