1. Понятие об инфекционном процессе. Механизмы, пути и факторы передачи инфекции. Входные ворота инфекции
 Скачать 0.68 Mb. Скачать 0.68 Mb.
|

|
16. Понятие об антигенах, их строение и свойства. Антигены бактерий и вирусов. Антигенами называют чужеродные для организма вещества коллоидной структуры, которые при попадании в его внутреннюю среду способны вызывать ответную специфическую иммунологическую реакцию, проявляющуюся, в частности, в образовании специфических антител, появлении сенсибилизированных лимфоцитов или в возникновении состояния толерантности к этому веществу. Вещества, являющиеся антигенами, должны быть чужеродны для организма, макромолекулярны, находиться в коллоидном состоянии, поступать в организм парентерально, т.е. минуя желудочно-кишечный тракт, в котором обычно происходит расщепление вещества и потеря его чужеродности. Под чужеродностью антигенов следует понимать определенную степень химического различия между антигеном и макромолекулами организма, во внутреннюю среду которого, но попадает. Антигенные свойства связаны с величиной молекулярной массы макромолекулы. Чем выше молекулярная масса вещества, тем выше его антигенность. Вместе с тем неверно считать, что высокая молекулярная масса является обязательным свойством антигена. Так, глюкогон, вазопрессин – ангиотензин также обладают антигенными свойствами. Принято различать полноценные антигены, неполноценные антигены (гаптены) и полугаптены. Полноценными антигенами называют такие, которые вызывают образование антител или сенсибилизацию лимфоцитов и способны реагировать с ними как в организме, так и в лабораторных реакциях. Свойствами полноценных антигенов обладают белки, полисахариды, высокомолекулярные нуклеиновые кислоты и комплексные соединения этих веществ. Неполноценные антигены, или гаптены, сами по себе не способны вызывать образование антител или сенсибилизацию лимфоцитов. Это свойство появляется лишь при добавлении к ним полноценных антигенов («проводников»), а среди образующихся антител или сенсибилизированных лимфоцитов часть специфична к «проводнику», а часть – к гаптену. Полугаптенами называют сравнительно простые вещества, которые при поступлении во внутреннюю среду организма могут химически соединяться с белками этого организма и придавать им свойства антигенов. К этим веществам могут принадлежать и некоторые лекарственные препараты (йод, бром, антипирин и др.). Молекула антигена состоит из двух неравных частей. Активная (малая часть) с носит название антигенной детерминанты (эпитоп) и определяет антигенную специфичность. Антигенные детерминанты расположены в тех местах молекулы антигена, которые находятся в наибольшей связи с микроокружением. В белковой молекуле, например, они могут располагаться не только на концах полипептидной цепи, но и в других ее частях. Антигенные детерминанты содержат в своем составе по крайней мере три аминокислоты с жесткой структурой (тирозин, триптофан, фенилаланин). Специфичность антигена связана также с порядком чередования аминокислот полипептидной цепи и комбинацией их положений по отношению друг к другу. Количество антигенных детерминант у молекулы антигена определяет его валентность. Она тем выше, чем больше относительная молекулярная масса молекулы антигена. Остальная (неактивная) часть молекулы антигена, как полагают, играет роль носителя детерминанты и способствует проникновению антигена во внутреннюю среду организма, его пиноцитозу или фагоцитозу, клеточной реакции на проникновение антигена, образование медиаторов межклеточного взаимодействия в иммунном ответе (Т-лимфоциты имеют рецепторы к носителю, В- к антигенной детерминанте). Соответственно анатомическим структурам бактериальной клетки различают Н-антигены (жгутиковые, если бактерия их имеет), К-антигены ( располагаются на поверхности клеточной сткнки), О-антигены (связан с клеточной стенкой бактерий), антигены экскретируемые бактериями в окружающую их среду (белки-экзотоксины, полисахариды капсул). Среди многочисленных антигенов микробной клетки различают такие, которые присущи только данному типу микробов (типовые антигены), данному виду (видовые антигены), а также общие для группы (семейства) микроорганизмов (групповые антигены). Таким образом, бактериальная клетка (как и микроорганизмы других царств микробов – вирусы, простейшие, грибки) представляют собой сложный комплекс многочисленных антигенов. При ее попадании во внутреннюю среду макроорганизма на многие из этих антигенов будут образовываться свои специфические антитела. Одни антигены индуцируют образование едва заметного количества антител (титр), другие – быстрое и значительное антителообразование. Соответственно этому различают «слабые» и «сильные» антигены. Не все антигены бактериальной клетки в равной степени участвуют в индукции невосприимчивости (иммунитета) к повторному попаданию в макроорганизм патогенных микробов того же вида. Способность антигена индуцировать иммунитет называют иммуногенностью, а такой антиген – иммуногеном. Установлено также, что определенные антигены некоторых микроорганизмов могут вызывать развитие различных типов гиперчувствительности (аллергии). Такие антигены называют аллергенами. По структуре вирусной чстицы различают несколько групп антигенов: ядерные, капсидные и суперкапсидные. Антигенный состав вириона зависит от строения самой вирусной частицы. Антигенная специфичность простоорганизованных вирусов связана с рибо- и дезоксинуклеопротеинами. У сложноорганизованных вирусов часть антигена связана с нуклеокапсидом, а другая локализуется во внешней оболочке – суперкапсиде. 17. Антитела (иммуноглобулины), структура, классы, функции. Понятие о моноклональных антителах. Гибридомы, получение, применение. Антитела (иммуноглобулины, ИГ, Ig) — особый класс гликопротеинов, присутствующих на поверхности B-лимфоцитов в виде мембраносвязанных рецепторов и в сыворотке крови и тканевой жидкости в виде растворимых молекул, и обладающих способностью очень избирательно связываться с конкретными видами молекул, которые в связи с этим называютантигенами. Антитела являются важнейшим фактором специфического гуморального иммунитета. Антитела используются иммунной системой для идентификации и нейтрализации чужеродных объектов — например, бактерий и вирусов. Антитела выполняют две функции: антиген-связывающую и эффекторную (вызывают тот или иной иммунный ответ, например, запускают классическую схему активации комплемента). Антитела синтезируются плазматическими клетками, которыми становятся некоторые В-лимфоциты, в ответ на присутствие антигенов. Для каждого антигена формируются соответствующие ему специализировавшиеся плазматические клетки, вырабатывающие специфичные для этого антигена антитела. Антитела распознают антигены, связываясь с определённым эпитопом — характерным фрагментом поверхности или линейной аминокислотной цепи антигена. Антитела представляют собой белки глобулиновой природы (иммуноглобулины) образующиеся в организме под воздействием антигена и обладающие способностью избирательно связываться с ним. Существуют пять разновидностей молекул (классов) иммуноглобулинов с молекулярной массой от 150 до 900 тыс. дальтон: IgM, lgG, IgA, IgE, IgD. Молекулы иммуноглобулинов состоят из двух легких (L) и двух тяжелых (Н) полипептидных цепей, соединенных между собой дисульфидными связями . Оба типа цепей, соединенных между собой, обладают антигенностью. У тяжелых цепей она специфична для каждого класса иммуноглобулинов и соответственно классам Н-цепи обозначаются m , g , a , e , s . Легкие цепи в антигенном отношении делятся на две разновидности - X и l , одинаковые для, разных классов. Антигенные различия тяжелых цепей используют для получения антисывороток, позволяющих выявить наличие в исследуемом материале иммуноглобулинов того или иного класса. Легкие цепи IgG состоят из двух участков (доменов): вариабельных (VL) и константных (CL). Тяжелые цепи включают в себя один вариабельный ( V Н) и 3 константных участка (CH 1 , CH 2 , СН 3 ). Вариабельные участки легких и тяжелых цепей формируют активные центры антител (VL -VH). Участок CL - CH 1 определяет небольшие различия в последовательности расположения аминокислот у индивидуумов одного и того же вида (аллоантигенные различия молекул IgM). Область CH 2 -CH 2 участвует в фиксации и активации комплемента, а область СН 3 -СН 3 - в фиксации антитела к клеткам (лимфоцитам, макрофагам, тучным клеткам). Данный тип строения молекулы характерен и для всех остальных классов иммуноглобулинов, различия заключаются в дополнительной организации этой основной единицы. Так, Н-цепь IgM состоит не из 4, а из 5 доменов, а вся молекула IgM представляет собой пентамер молекулы IgG, соединенный дополнительными полипептидными J-цепями. IgA может быть в форме мономеров, димеров и секреторного IgA. Последние две формы имеют дополнительные (димеры) J или J и S цепи (секреторный). Другие свойства антител представлены в таблице 5. Таблица 5. Основные характеристики иммуноглобулинов человека
Молекула антитела связывается с детерминантой антигена не целиком, а лишь определенной своей частью, называемой активным центром. Активный центр представляет собой полость или щель, соответствующую пространственной конфигурации детерминантной группы антигена. Один из активных центров по разным причинам может быть функционально инертным. Такие антитела называются неполными. Их появлению обычно предшествует образование полных, т. е. антител с двумя (IgG) активными центрами. Неполные антитела встречаются у разных классов иммуноглобулинов. Основная масса антител образуется в клетках плазмоцитарного ряда (плазмобласт, проплазмоцит, плазмоцит). Каждая из них продуцирует антитела только одной специфичности, т. е. к одной антигенной детерминанте. Территориально эти клетки располагаются в селезенке, лимфоузлах, костном мозге, лимфоидных образованиях слизистых оболочек. При первичном контакте организма с антигеном и антителообразовании различают индуктивную и продуктивные фазы. Продолжительность первой фазы составляет около 2 суток. В этот период происходит пролиферация и дифференцировка лимфоидных клеток, развитие плазмобластической реакции. Вслед за индуктивной наступает продуктивная фаза. В сыворотке крови антитела начинают определяться с З-го дня после контакта с антигеном. Эти антитела относятся к классу IgM. С 5-7 дня происходит постепенная смена синтеза IgM на синтез IgG той же специфичности. Обычно к 12-15 дню кривая антителообразования достигает максимума, далее уровень антител начинает снижаться, но определенное их количество можно обнаружить и через много месяцев, а иногда и лет. При повторном контакте организма с тем же антигеном индуктивная фаза занимает лишь несколько часов. Продуктивная фаза протекает быстрее и интенсивнее, осуществляется синтез преимущественно IgG. 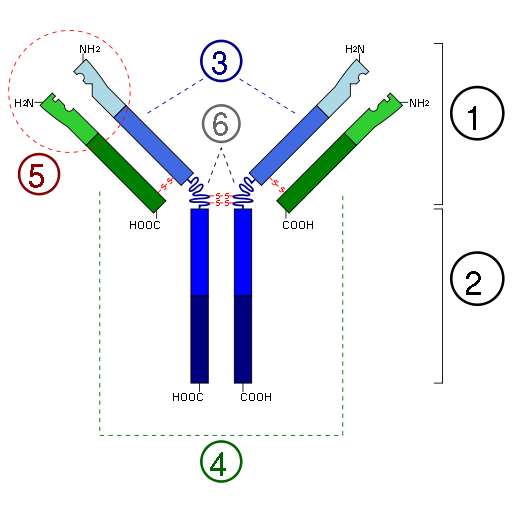 Иммуноглобулины всех изотипов бифункциональны. Это означает, что иммуноглобулин любого типараспознает и связывает антиген, а затем усиливает киллинг и/или удаление иммунных комплексов, сформированных в результате активации эффекторных механизмов. Одна область молекулы антител (Fab) определяет её антигенную специфичность, а другая (Fc) осуществляет эффекторные функции: связывание с рецепторами, которые экспрессированы на клетках организма (например, фагоцитах); связывание с первым компонентом (C1q) системы комплемента для инициации классического пути каскада комплемента.
Классификация по антигенам
|