колоквиум №2. 1. Предмет, задачи, методы генетики. История развития генетики. Роль отечественных ученых (Н. К. Кольцов, А. С. Серебровский, С. С. Четвериков) в развитии генетики
 Скачать 274.6 Kb. Скачать 274.6 Kb.
|

|
10 Хромосомная теория наследственности. Сцепление генов. Кроссинговер. Метод соматической гибридизации клеток, его применение при картировании генов человека в хромосомах Положения хромосомной теории наследственности: 1 Гены локализованы в хромосомах. линейно в определенных локусах 2. Аллельные гены занимают одинаковые локусы в гомологичных хромосомах. 3. При этом различные хромосомы содержат неодинаковое число генов 4 Кроме того, набор генов каждой из негомологичных хромосом уникален 5. Гены одной хромосомы образуют группу сцепления, благодаря нему происходит сцепленное наследование некоторых признаков. число их равно гаплоидному набору хромосом. При этом сила сцепления находится в обратной зависимости от расстояния между генами. 6 Каждый биологический вид характеризуется определенным набором хромосом — кариотипом. 7. Между гомологичными хромосомами возможен обмен аллельными генами (кроссинговер). 8. Расстояние между генами пропорционально проценту кроссинговера между ними и выражается в морганидах (1 морганида равна 1% кроссинговера). Группа сцепления — хромосома, в которой расположено большое число генов. Соответствие групп сцепления числу хромосом. Гены, локализованные в одной хромосоме, наз-я группой сцепления. Кроссинговер (от англ. crossingover), перекрест, взаимный обмен участками парных хромосом, происходящий в результате разрыва и соединения в новом порядке их нитей — хроматид (рис.); приводит к перераспределению (рекомбинации) сцепленных генов. Т. о., К. — важнейший механизм, обеспечивающий комбинаторную изменчивость, а следовательно, — один из главных. факторов эволюции. К., как правило, имеет место в профазе первого деления половых клеток (см. Мейоз), когда их хромосомы представлены, четырьмя нитями. Соматические клетки содержат весь объем генетической информации. Это дает возможность изучать многие вопросы генетики человека, которые невозможно исследовать на целом организме. Благодаря методам генетики соматических клеток человек как бы стал одним из экспериментальных объектов. Чаще всего используют клетки соединительной ткани (фибробласты) и лимфоциты крови. культивирование клеток вне организма позволяет получить достаточное количество материала для исследования. что не всегда возможно взять у человека без ущерба для здоровья. Находящиеся в культуре клетки какой-либо ткани можно подвергать изучению различными методами: цитологическим, биохимическим, иммунологическим. такое исследование может быть в ряде случаев более точным, чем на уровне целостного организма, так как метаболические процессы удается выделить из сложной цепи взаимосвязанных реакций. происходящих в организме. В 1960 г. французский биолог Ж. Барский, выращивая вне организма в культуре ткани клетки двух линий мышей, обнаружил, что некоторые клетки по своим морфологическим и биохимическим признакам были промежуточными между исходными родительскими клетками. Эти клетки оказались гибридными. Такое спонтанное слияние клеток в культуре ткани происходит довольно редко. В дальнейшем оказалось, что частота гибридизации соматических клеток повышается при введении в культуру клеток РНК-содержащего вируса парагриппа Сендай, который как вообще все вирусы, изменяет свойства клеточных мембран и делает возможным слияние клеток. Под влиянием такого вируса в смешанной культуре двух типов клеток образуются клетки, содержащие в общей цитоплазме ядра обеих родительских клеток – гетерокарионы. После митоза и последующего разделения цитоплазмы из двуядерного гетерокариона образуются две одноядерные клетки, каждая из которых представляет собой синкарион – настоящую гибридную клетку, имеющую хромосомы обеих родительских клеток. месте с тем сначала можно было картировать только гены человека, проявляющиеся на клеточном уровне, и невозможно было работать с генами, имевшими сложное феноти-пическое проявление, при котором первичный дефект оставался неизвестным. Методы молекулярной генетики позволили решить эту проблему, и в настоящее время с помощью ПЦР, блоттинга по Саузерну и иных молекулярно-генетиче-ских методов можно тестировать гибридные соматические клетки на наличие в оставшихся хромосомах человека любых генов независимо от того, известен ли продукт этих генов или нет. Недостатком метода картирования генов человека с помощью гибридных соматических клеток было то, что часто локализация генов устанавливалась с точностью до хромосомы. Разрешающие возможности в картировании генов таким путем были, следовательно, ограничены. 11Наследование. Типы наследования. Особенности аутосомного, Х-сцепленного и голандрического типов наследования. Полигенное наследование наследственностью - свойство клеток или организмов в процессе самовоспроизведения передавать новому поколению способность к определенному типу обмена веществ и индивидуального развития, в ходе которого у них формируются общие признаки и свойства данного типа клеток и вида организмов, а также некоторые индивидуальные особенности родителей. На популяционно-видовом уровне организации жизни наследственность проявляется в поддержании постоянного соотношения различных генетических форм в ряду поколений организмов данной популяции (вида). Наследственность – свойство живых организмов, обеспечивающее материальную преемственность онтогенеза в определенных условиях внешней среды. Гены детерминируют последовательность полипептидной цепи. Наследование – передача информации от одного поколения к другому. Благодаря наследственности стало возможно существование популяций, видов и других групп. 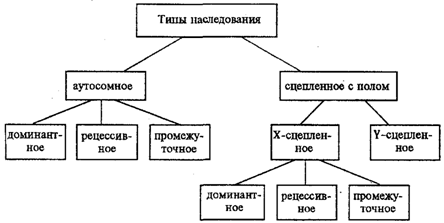 Аутосомное наследование. Характерные черты аутосомного наследования признаков обусловлены тем, что соответствующие гены, расположенные в аутосомах, представлены у всех особей вида в двойном наборе. Это означает, что любой организм получает такие гены от обоих родителей. В соответствии с законом чистоты гамет в ходе гаметогенеза все половые клетки получают по одному гену из каждой аллельной пары (рис. 6.6). Обоснованием этого закона является расхождение гомологичных хромосом, в которых располагаются аллельные гены, к разным полюсам клетки в анафазе I мейоза Ввиду того что развитие признака у особи зависит в первую очередь от взаимодействия аллельных генов, разные его варианты, определяемые разными аллелями соответствующего гена, могут наследоваться по аутосомно-доминантному или аутосомно-рецессивному типу, если имеет место доминирование. Возможен также промежуточный тип наследования признаков при других видах взаимодействия аллелей (см. разд. 3.6.5.2). При доминировании признака, описанном Г. Менделем в его опытах на горохе, потомки от скрещивания двух гомозиготных родителей, различающихся по доминантному и рецессивному вариантам данного признака, одинаковы и похожи на одного из них (закон единообразия F1). Описанное Менделем расщепление по фенотипу в F2 в отношении 3:1 в действительности имеет место лишь при полном доминировании одного аллеля над другим, когда гетерозиготы фенотипически сходны с доминантными гомозиготами (закон расщепления в F2). Наследование рецессивного варианта признака характеризуется тем, что он не проявляется у гибридов F1, а в F2 проявляется у четверти потомков. В случаях формирования у гетерозигот нового варианта признака по сравнению с гомозиготами, что наблюдается при таких видах взаимодействия аллельных генов, как неполное доминирование, кодоминирование, межаллельная комплементация, гибриды F1 не похожи на родителей, а в F2 образуется три фенотипических группы потомков (рис. 6.7, II). Сцепленное с полом наследование. Х-сцепленное наследование. Х-хромосома присутствует в кариотипе каждой особи, поэтому признаки, определяемые генами этой хромосомы, формируются у представителей как женского, так и мужского пола. Особи гомогаметного пола получают эти гены от обоих родителей и через свои гаметы передают их всем потомкам. Представители гетерогаметного пола получают единственную Х-хромосому от гомогаметного родителя и передают ее своему гомогаметному потомству. У млекопитающих (в том числе и человека) мужской пол получает Х-сцепленные гены от матери и передает их дочерям. При этом мужской пол никогда не наследует отцовского Х-сцепленного признака и не передает его своим сыновьям. Так как у гомогаметного пола признак развивается в результате взаимодействия аллельных генов, различают Х-сцепленное доминантное и Х-сцепленное рецессивное наследование. Х-сцепленный доминантный признак (красный цвет глаз у дрозофилы) передается самкой всему потомству. Самец передает свой Х-сцепленный доминантный признак лишь самкам следующего поколения. Самки могут наследовать такой признак от обоих родителей, а самцы — только от матери. Х-сцепленный рецессивный признак, (белый цвет глаз у дрозофилы) у самок проявляется только при получении ими соответствующего аллеля от обоих родителей (XaXa). У самцов XaY он развивается при получении рецессивного аллеля от матери. Рецессивные самки передают рецессивный аллель потомкам любого пола, а рецессивные самцы —только «дочерям» (см. рис. 6.8). Голандрическое наследование. Активно функционирующие гены Y-хромосомы, не имеющие аллелей в Х-хромосоме, присутствуют в генотипе только гетерогаметного пола, причем в гемизиготном состоянии. Поэтому они проявляются фенотипически и передаются из поколения в поколение лишь у представителей гетерогаметного пола. Так, у человека признак гипертрихоза ушной раковины («волосатые уши») наблюдается исключительно у мужчин и наследуется от отца к сыну. Большинство количественных признаков организмов определяется полигенами, т. е. системой неаллельных генов, одинаково влияющих на формирование данного признака. Взаимодействие таких генов в процессе формирования признака называют полимерным. Оно сводится чаще всего к суммированию действия сходных аллелей этих генов, определяющих формирование одинакового варианта признака. Совместное действие полигенов обусловливает различную экспрессивность — степень выраженности признака, зависящую от дозы соответствующих аллелей. В основе появления в геноме таких генов, очевидно, лежит их дупликация или амплификация (см. разд. 3.6.4.3). Это позволяет увеличить синтез соответствующего продукта в клетках организма. По полимерному типу взаимодействия у человека определяется интенсивность окраски кожных покровов, зависящая от уровня отложения в клетках пигмента меланина. В геноме человека имеется четыре гена, отвечающих за этот признак. В генотипе все они представлены в двойной дозе. В связи с тем что эти гены отвечают за развитие одного и того же признака, их обозначают одной и той же буквой, но с разными символами, чтобы подчеркнуть их неаллельность — P1P2P3P4. Наличие в генотипе восьми доминантных аллелей в системе полигенов, определяющих цвет кожи, обусловливает максимальную ее пигментацию, наблюдаемую у африканских негров (P1P1P2P2P3P3P4P4). Полное отсутствие доминантных аллелей у. рецессивных гомозигот (р1р2р2р3р3р4р4) проявляется в виде минимальной пигментации у европеоидов. Большее или меньшее количество доминантных аллелей, колеблющееся от 8 до 0, обеспечивает разную интенсивность окраски кожи (рис. 3.80). Полимерное взаимодействие генов лежит в основе определения главным образом количественных признаков (рост, масса организма, возможно, интеллект). полигенное наследование было обнаружено у гималайских кроликов в степени пегости. Гомозиготные черные особи имеют белые пятна только около глаз и на носу, а гомозиготные белые имеют черные пятна на кончиках лап и в основании хвоста. При их скрещивании образовывались особи с промежуточной окраской, а в F2 имела место формула 1:4:6:4:1. При тригибридном скрещивании формула расщепления будет выглядеть так 1:6:15:20:15:6:1. Явление полимерии у человека наблюдается в степени окраски кожи. От браков негров с альбиносами рождаются дети с промежуточной окраской кожи (мулаты). Дети, родители которых мулаты, могут иметь различную окраску кожи от белой до темной. Считают, что на степень окраски кожи влияют 3 или 4 полигена... На полигенное наследование немаловажное влияние оказывают негенетические факторы, такие как климат, питание, болезни. Причем чем больше полигенов влияют на проявление признака, тем он устойчивее по отношению к негенетическим факторам. Большое значение при полимерии занимает трансгрессия. При трансгрессии в поколении F2 половина особей будет иметь более выраженный признак, чем у родительских форм, а другая половина менее выраженный признак. Явление трансгрессии используется в селекционной деятельности. При этом искусственный отбор будет направлен на закрепление признаков у особей с положительной трансгрессией (Al A1A2A2...) и удаление особей с отрицательной трансгрессией (alala2a2...). 12Ген, его свойства. Генетический код, его свойства. Структура и виды РНК. Ген-участок ДНК, который несет в себе информацию о строении какого либо белка Свойства: 1 дискретность действия- развитие различных признаков контролируется разными генами. 2 стабильность - передается в ряду поколений в неизменном виде. 3 специфичность - каждый из генов обуславливает развитие определенного признака. 4 плейотропия - способность генов обеспечивать развитие одновременно нескольких признаков Классификация генов 1)По характеру взаимодействия в аллельной паре: - доминантный (ген, способный подавлять проявление аллельного ему рецессивного гена); -рецессивный (ген, проявление которого подавлено аллельным ему доминантным геном). 2) Функциональная классификация . Генетический код и его свойства-свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов: 1)Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон). 2)Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно. 3)Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки). 4)Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте 5)Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов. 6)Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека. 7)Помехоустойчивость — мутации замен нуклеотидов не приводящие к смене класса кодируемой аминокислоты, называют консервативными; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными. Структурные :кодирующие белки кодирующие т-РНК, кодирующие р-РНК. Регуляторные: гены ингибиторы (подавляют активность генов), гены интерсификаторы (повышают активность генов), гены репараторы (исправляющие мутации) 13.Процессинг, сплайсинг. Роль РНК в процессе реализации наследственной информации Процессинг РНК (посттранскрипционные модификации РНК) — совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта РНК в зрелую РНК. Наиболее известен процессинг матричных РНК, которые во время своего синтеза подвергаются модификациям: кэпированию, сплайсингу и полиаденилированию. Также модифицируются (другими механизмами) рибосомные РНК, транспортные РНК и малые ядерные РНК. Сплайсинг (от англ. splice — сращивать или склеивать концы чего-либо) — процесс вырезания определенных нуклеотидных последовательностей из молекул РНК и соединения последовательностей, сохраняющихся в «зрелой» молекуле, в ходе процессинга РНК. Наиболее часто этот процесс встречается при созревании информационной РНК (мРНК) у эукариот, при этом путём биохимических реакций с участием РНК и белков из мРНК удаляются участки, не кодирующие белок (интроны) и соединяются друг с другом кодирующие аминокислотную последовательность участки — экзоны. Таким образом незрелая пре-мРНК превращается в зрелую мРНК, с которой считываются (транслируются) белки клетки. Большинство генов прокариот, кодирующих белки, не имеют интронов, поэтому у них сплайсинг пре-мРНК встречается редко. У представителей эукариот, бактерий и архей встречается также сплайсинг транспортных РНК (тРНК)[1] и других некодирующих РНК. Процессинг и сплайсинг способны объединять структуры, удаленные друг от друга, в один ген, поэтому они имеют огромное эволюционное значение. Подобные процессы упрощают видообразование. Белки имеют блочную структуру. Например, фермент – ДНК-полимераза. Он представляет собой непрерывную полипептидную цепь. Он состоит из собственной ДНК-полимеразы и эндонуклеазы, которая расщепляет молекулу ДНК с конца. Фермент состоит из 2 доменов, которые образуют 2 независимые компактные частицы, связанные полипептидным мостиком. На границе между 2мя генами ферментов находится интрон. Когда-то домены были раздельными генами, а затем – сблизились. Нарушения подобной структуры гена приводит к генным болезням. Нарушение строения интрона фенотипически незаметно, нарушение в экзонной последовательности приводят к мутации (мутации глобиновых генов). Биосинтез белка — сложный многостадийный процесс синтеза полипептидной цепи из аминокислотных остатков, происходящий на рибосомах клеток живых организмов с участием молекул мРНК и тРНК. Биосинтез белка можно разделить на стадии транскрипции, процессинга и трансляции. Во время транскрипции происходит считывание генетической информации, зашифрованной в молекулах ДНК, и запись этой информации в молекулы иРНК. В ходе ряда последовательных стадий процессинга из мРНК удаляются некоторые фрагменты, ненужные в последующих стадиях, и происходит редактирование нуклеотидных последовательностей. После транспортировки кода из ядра к рибосомам происходит собственно синтез белковых молекул, путём присоединения отдельных аминокислотных остатков к растущей полипептидной цепи. Роль посредника, функцией которого является перевод наследственной информации, сохраняемой в ДНК, в рабочую форму, играют рибонуклеиновые кислоты - РНК. рибонуклеиновые кислоты представлены одной полинуклеотидной цепью, которая состоит из четырех разновидностей нуклеотидов, содержащих сахар, рибозу, фосфат и одно из четырех азотистых оснований - аденин, гуанин, урацил или цитозин Матричная, или информационная, РНК (мРНК, или иРНК). Транскрипция. Для того чтобы синтезировать белки с заданными свойствами, к месту их построения поступает "инструкция" о порядке включения аминокислот в пептидную цепь. Эта инструкция заключена в нуклеотидной последовательности матричных, или информационных РНК (мРНК, иРНК), синтезируемых на соответствующих участках ДНК. Процесс синтеза мРНК называют транскрипцией. В процессе синтеза, по мере продвижения РНК-полимеразы вдоль молекулы ДНК, пройденные ею одноцепочечные участки ДНК вновь объединяются в двойную спираль. Образуемая в ходе транскрипции мРНК содержит точную копию информации, записанной в соответствующем участке ДНК. Тройки рядом стоящих нуклеотидов мРНК, шифрующие аминокислоты, называют кодонами. Последовательность кодонов мРНК шифрует последовательность аминокислот в пептидной цепи. Кодонам мРНК соответствуют определенные аминокислоты (табл.1). Транспортная РНК (тРНК). Трансляция. Важная роль в процессе использования наследственной информации клеткой принадлежит транспортной РНК (тРНК). Доставляя необходимые аминокислоты к месту сборки пептидных цепей, тРНК выполняет функцию трансляционного посредника. В ней выделяют четыре главные части, выполняющие различные функции. Акцепторный "стебель" образуется двумя комплементарно соединенными концевыми частями тРНК. Он состоит из семи пар оснований.3'-конец этого стебля несколько длиннее и формирует одноцепочечный участок, который заканчивается последовательностью ЦЦА со свободной ОН-группой. К этому концу присоединяется транспортируемая аминокислота. Остальные три ветви представляют собой комплементарно спаренные последовательности нуклеотидов, которые заканчиваются неспаренными участками, образующими петли. Средняя из этих ветвей - антикодоновая - состоит из пяти пар нуклеотидов и содержит в центре своей петли антикодон. Антикодон - это три нуклеотида, комплементарные кодону мРНК, который шифрует аминокислоту, транспортируемую данной тРНК к месту синтеза пептида. В целом различные виды тРНК характеризуются определенным постоянством нуклеотидной последовательности, которая чаще всего состоит из 76 нуклеотидов. Варьирование их числа связано главным образом с изменением количества нуклеотидов в дополнительной петле. Комплементарные участки, поддерживающие структуру тРНК, как правило, консервативны. Первичная структура тРНК, определяемая последовательностью нуклеотидов, формирует вторичную структуру тРНК, имеющую форму листа клевера. В свою очередь, вторичная структура обусловливает трехмерную третичную структуру, для которой характерно образование двух перпендикулярно расположенных двойных спиралей (рис.27). Одна из них образована акцепторной и ТψС-ветвями, другая - антикодоновой и D-ветвями. На конце одной из двойных спиралей располагается транспортируемая аминокислота, на конце другой - антикодон. Эти участки оказываются максимально удаленными друг от друга. Стабильность третичной структуры тРНК поддерживается благодаря возникновению дополнительных водородных связей между основаниями полинуклеотидной цепи, находящимися в разных ее участках, но пространственно сближенных в третичной структуре. Различные виды тРНК имеют сходную третичную структуру, хотя и с некоторымивариациями. Одной из особенностей тРНК является наличие в ней необычных оснований, возникающих вследствие химической модификации уже после включения нормального основания в полинуклеотидную цепь. Эти измененные основания обусловливают большое структурное многообразие тРНК при общем плане их строения. |